
トマト黄化壊疽ウイルス
【課題】異なったトマト黄化壊疽ウイルス(TSWV)単離株による感染に抵抗するトランスジェニック植物を作出すること。
【解決手段】TSWV-B S(ブラジリアン単離株)のRNAをクローニングし、該TSWV-Bを特徴付けする。TSMV単離株からの該ヌクレオキャプシドのヌクレオチド配列を含有するトランスジェニック植物は、異なる血清群からのトスポウイルスに対する抵抗性を付与する。加えて、TSWVのレタス単離株からのヌクレオキャプシドのヌクレオチド配列を含有するトランスジェニック植物を作出し、それが同種および近縁双方のウイルス単離株に対する抵抗性を当該トランスジェニック植物で生じる(少量のヌクレオキャプシド蛋白質を産生する植物において)のに対して、多量のヌクレオキャプシド蛋白質を産生する植物は、同種単離株および遠縁のツリフネソウ壊疽斑点ウイルス(INSV)の単離株双方に対して中レベルの保護を有していた。
【解決手段】TSWV-B S(ブラジリアン単離株)のRNAをクローニングし、該TSWV-Bを特徴付けする。TSMV単離株からの該ヌクレオキャプシドのヌクレオチド配列を含有するトランスジェニック植物は、異なる血清群からのトスポウイルスに対する抵抗性を付与する。加えて、TSWVのレタス単離株からのヌクレオキャプシドのヌクレオチド配列を含有するトランスジェニック植物を作出し、それが同種および近縁双方のウイルス単離株に対する抵抗性を当該トランスジェニック植物で生じる(少量のヌクレオキャプシド蛋白質を産生する植物において)のに対して、多量のヌクレオキャプシド蛋白質を産生する植物は、同種単離株および遠縁のツリフネソウ壊疽斑点ウイルス(INSV)の単離株双方に対して中レベルの保護を有していた。
【発明の詳細な説明】
【技術分野】
【0001】
トスポウイルス属のウイルスは広範な植物種、特に、タバコ、ラッカセイ、蔬菜および観賞植物に感染する。2種のウイルス種、トマト黄化壊疽ウイルス(TSWV)およびツリフネソウ属壊疽斑点ウイルス(INSV)は、トスポウイルス属に属すると認識されている。
【背景技術】
【0002】
トマト黄化壊疽ウイルス(TSWV)は、核酸-蛋白質複合体がリポ蛋白質エンベロープによって覆われており、かつ唯一のアザミウマ媒介ウイルスである点において植物ウイルスの中では特異である。最近、このウイルスは、ブニヤウイルス科のトスポウイルス属に分類された。TSWVウイルス粒子には、29Kのヌクレオキャプシド蛋白質(「NP」または「N」)、2種の膜関連糖蛋白質(58Kおよび78K)、ならびに恐らくウイルス逆転写酵素のための大きな200K蛋白質が含まれる[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:2207頁(1991年);バイロロジー(Virol.)第56:12頁(1973年);およびジャーナル・ジェネラル・バイロロジー(J.Gen.Virol.)第36巻:267頁(1977年)参照]。そのウイルスゲノムは、L RNA(8900ヌクレオチド)、M RNA(5400ヌクレオチド)およびS RNA(2900ヌクレオチド))と命名される3本の負鎖(-)RNAよりなり[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第36巻:81頁(1977年);ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第53巻:12頁(1981年);およびジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第70巻:3469頁(1989年)参照]、その各々はNPによって包まれている。3種のTSWV単離株からのS RNAの部分配列または全長配列によって、両側センス(ambisense)遺伝子配置を有する2つのオープンリーディングフレーム(ORF)の存在が明らかとなった[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:1頁(1990年)およびジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第72巻:461頁(1991年)参照]。大きな方のオープンリーディングフレームはウイルスRNA鎖上に位置し、52Kの非構造蛋白質をコードする余地を有している。小さな方のORFはウイルスの相補的RNA鎖上に位置し、サブゲノムRNAを通して29KのNPに翻訳される。
【0003】
58Kおよび78Kの膜関連糖蛋白質をコードするオープンリーディングフレームでの両側センス鎖暗号化戦略も、TSWV M RNAに特徴的である。TSWV L RNAは、恐らくはウイルス逆転写酵素のための大きな200K蛋白質をコードすることが配列決定されている。
【0004】
2種のTSWV血清群である「L」および「I」が、構造蛋白質の血清学的分析および細胞病理学的な構造の形態学に基づいて同定され特徴付けられている[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:933頁(1990年)およびファイトパソロジー(Phytopathology)第81巻:525頁(1991年)参照]。それらは、血清学的に保存されたG1およびG2糖蛋白質を有しているが、「I」血清群のNPは「L」血清群のものとは血清学的に異なる。「L」および「I」血清群の間でNPを比較した結果、ヌクレオチドおよびアミノ酸のレベルで各々62%および67%の同一性が示された(ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第72巻:2597頁(1991年)参照]。
【0005】
TSWVは広範な宿主範囲を有し、50科の360植物種を超える植物に感染して、蔬菜および観賞植物に重大な経済的損失を全世界的に引き起こしている。「L」血清群が蔬菜およびタバコのごとき農作物に幅広く認められる一方、「I」血清群は観賞作物にたいてい限定されている。スイカおよび他のウリ科植物に全身感染し、かつそのNPはいずれの血清群のものとも血清学的に関連性がないために、区別される単離株としてウリ科植物単離株が最近同定された[プラント・ディジーズ(Plant Disease)第68巻:1006頁(1984年)参照]。抵抗性植物の育種または非遺伝的方法によってTSWV病の拡大を低下させることが時々できることもあるが、これらの慣用的方法による病害の完全な制御は一般的に困難であることが証明されている[プラント・ディジーズ(Plant Disease)第73巻:375頁(1989年)]。
【発明の開示】
【発明が解決しようとする課題】
【0006】
1986年以来、ウイルスの外皮蛋白質(CP)遺伝子を導入したトランスジェニック植物が該ウイルスによる感染に対してしばしば抵抗性を示すことを、膨大な報告が示してきた。この現象は、通常、外皮蛋白質-媒介保護(CPMP)と呼ばれている。保護の程度は、病徴発現の遅延から病徴およびウイルス蓄積の欠如までに及ぶ。最近の2報の独立した報告[バイオル・テクノロジー(Biol.Tech-nology)第9巻:1363頁(1991年)およびモレキュラー・アンド・プラント-マイクローブ・インタラクションズ(Mol.Plant-Microbe Interact.)第5巻:34頁(1992年)]は、TSWVのヌクレオキャプシド蛋白質(NP)遺伝子を発現するトランスジェニック・タバコ植物が同種の単離株による感染に対して抵抗性を示すことを示した。しかしながら、TSWVは多種の生物的に多様な株に拡散しているため、異なったTSWV単離株による感染に抵抗するトランスジェニック植物の有効性を試験することが非常に重要である。本発明の知見は、本発明によるトランスジェニック植物が、「L」血清群の2種の異種単離株および「I」血清群の1種の単離株に対して抵抗性を示したことを証明することによって、先行の報告のものを超えるものである。本発明者らは、高レベルのNPを蓄積するトランスジェニック植物が「I」血清群の単離株に抵抗性を示す一方、たとえあるとしても非常に低いレベルNPしか蓄積していない植物において、「L」血清群の2種の異種単離株に対する抵抗性が主に認められたことも示す。
【0007】
しかしながら、高レベルのNP蛋白質を蓄積する植物が病徴発現の遅延を示したにも拘わらず、ブラジリアン(Brazilian)単離株に対する抵抗性は認められなかった。TSWV-Bと命名されたこのブラジリアン単離株は、「L」および「I」血清群とは血清学的に区別されるN蛋白質を有し、TSWV-Bがメロンおよびカボチャに全身感染しない点においてウリ科単離株とは生物学的に異なる。
【課題を解決するための手段】
【0008】
従って、本発明の1つの態様は、TSWV-B SのRNAをクローニングし、配列決定して、他のTSWV単離株の公表されている配列と比較することによって該TWV-Bを特徴付けすることである。
【発明を実施するための最良の形態】
【0009】
本発明の種々の態様は、以下の実施例、図面およびデータを包含する本発明の詳細な説明から容易に明らかとなるであろう。
図1は、本発明によるウイルスRNAからNP遺伝子をクローニングするための戦略を示す図である。
図2は、タバコ・プロトプラストにおける本発明によるトマト黄化壊疽病ウイルスのヌクレオキャプシド蛋白質(NP)遺伝子のイン・ビボ(in vivo)経時的発現を示す図である。
図3は、本発明によるTSWV-B S RNAにおける、配列決定したcDNAクローンの位置を示す図である。
図4は、本発明によるTSWV単離株の中での関係を示す樹系図である。
図5は、本明細書に記載したTSWV単離株の血清学的な関係を示す図である。
図6は、トランスジェニック植物におけるヌクレオキャプシド蛋白質(NP)の蓄積のレベルとTSWV単離株に対する抵抗性の度合との相関を示す図である。
図7は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BL N暗号配列を示す図である。
図8は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BLの半分のN遺伝子断片を示す図である。
【0010】
さらに詳細には、図2は、ポリエチレングリコール(PEG)を用いてタバコ葉肉プロトプラストに構築物が移入されたNP遺伝子の経時的発現を図示している。続いて、NP遺伝子を発現させるためにその形質転換プロトプラストを2日間インキュベートした。プロトプラストから蛋白質を抽出し、TSWV NPに対する抗体を用いた二重抗体サンドイッチ酵素結合免疫吸着アッセイ(DAS-ELISA)によってNPについて試験した。NP−およびNP+は、各々プラスミドpBI525-NP−およびpBI525-NP+で形質転換したプロトプラストを表す。コーティング用の抗体濃度は5μg/ml、酵素結合物の希釈率は1:250である。データは、基質添加後30、60および90分に収集した。
【0011】
図3において、5つの重複cDNAクローンは、TSWV-BのS RNA地図以下の尺度で示されている。これらのクローンは、TSWV-Bに感染したエヌ・ベンサミアーナ(N.benthamiana)から単離した二本鎖RNAからのランダムプライマーで合成した。
【0012】
図4において、配列は、GCGシークエンス・アナリシス・ソフトウェア・パッケージ(GCG Sequence analysis software package)のパイルアップ・プログラムを用いて比較した。水平線は遺伝子距離に比例するが、垂直線は任意の長さであり、有意性はない。
【0013】
さらに詳細には、図5において、エヌ・ベンサミアーナ・ドミン(N.benthamianaDomin)をTSWV単離株[TSWV-BL(レタス単離株)、アルカンサス(Arkansas)、10W体菜(pakchoy)(TSWV-10W))、ベゴニアおよびブラジル(TSWV-B)]で感染させた。感染リーフディスク(0.05g)を12mlの酵素結合緩衝液中にて摩砕し、TSWV-BLビリオン(BL ビリオン)、またはTSWV-BLのNP(BL-NP)もしくはTSWV-IのNP(I-NP)に対して生起させた抗体を用いるDAS-ELISAにより分析した。コーティング用の抗体濃度は1μg/ml;BLウイルス粒子については結合物の希釈率は1:2000、BL-NPについては希釈率は1:250、I-NPについては希釈率は1:1000であった。基質添加後10分(BL)、50分(BL-NP)または30分に結果を収集した。
【0014】
図6に関しては、TSWV-BLのNPに対して生起させた抗体でNP蓄積につきDAS-ELISAでトランスジェニック植物をアッセイした。植物を基質添加後150分に判読し、該トランスジェニック植物を4つのカテゴリーにグループ分けした:0.050より低いOD405nm値、0.050ないし0.200の間のOD405nm値、0.200ないし0.400の間のOD405nm値、および0.400よりも高いOD405nm値。対照NP(-)植物のOD405nm判読値は0ないし0.05であった。同一の植物をアルカンサス(Ark)および10W体菜(10W)単離株またはベゴニア単離株で攻撃し、各植物の感受性を接種約12日後に記録した。結果は、アルカンサスおよび10Wチュウゴクキャベツ単離株で接種した51個体のR1 NP(+)植物、およびベゴニア単離株で接種した139個体のR1 NP(+)植物から収集した。バー上の数字は、試験したR1 NP(+)植物の合計数を表す。
【0015】
実施例I
TSWV-BL RNAの単離:
TSWV-BL単離株は、ダチュラ・ストラモニウム・エル(Datura stramonium L.)から以下のごとく精製した:感染組織を3倍容量の緩衝液(0.033MのKH2PO4、0.067MのK2HPO4、0.01MのNa2SO3)と共にワーリング・ブレンダーで45秒間摩砕した。そのホモジネートを前記緩衝液で湿らせた4層のチーズクローズを通して濾過し、7,000rpmにて15分間遠心した。そのペレットを組織の元の重量に等しい量の0.01M Na2SO3に再懸濁し、8,000rpmにて15分間再度遠心した。その後に、上清を元の組織重量の1/10に等しい量の0.01M Na2SO3に再懸濁した。そのウイルス抽出液を9,000rpmにて15分間遠心し、その上清を、0.01MのNa2SO3中に作成した10-40%スクロース段階勾配上に注意深く負荷した。23,000rpmにて35分間遠心した後に、ウイルスゾーン(メニスカスより約3cm下)を採取し、2倍容量の0.01M Na2SO3で希釈した。その半-精製ウイルスを27,000rpmにて55分間ペレット化した。
【0016】
実施例II
TSWVおよびウイルスRNAの精製
TSWV-BL単離株[プラント・ディジーズ(Plant Disease)第74巻:154頁(1990年)]は、実施例Iに記載のごとく、ダチュラ・ストラモニウム・エル(Datura stramonium L)から精製した。0.04%のベントナイト、10μg/mlのプオテイナーゼK、0.1Mの炭酸アンモニウム、0.1%(w/v)のジエチルチオカルバミン酸ナトリウム、1mMのEDTA、および1%(w/v)のドデシル硫酸ナトリウム(SDS)の溶液中に精製したウイルスを再懸濁し、65℃にて5分間インキュベートして、直ちにH2O-飽和フェノールから抽出し、続いてさらにクロロホルム/イソアミルアルコール(24:1)から抽出した。ウイルスRNAを2.5容量のエタノール中に沈殿させ、蒸留水に溶解した。
【0017】
実施例III
cDNAおよびPCRに基づくNP遺伝子のクローニング:
第一鎖cDNAは、グブラー(Gubler)およびホフマン(Hoffman)により記載されているランダムプライマーを用いて、精製したTSWV-BLのRNAから合成した[ジーン(Gene)第25巻:263頁(1983年)参照]。第二鎖は、RNアーゼ H/DNAポリメラーゼで試料を処理することにより作製した。得られた二重鎖cDNA試料を、スクロース濃度勾配遠心法によりサイズ-分画し、EcoRIメチラーゼによりメチル化し、EcoRIリンカーを付加した。EcoRIで消化した後に、子ウシ腸アルカリ性ホスファターゼで処理することによってその5'-末端リン酸基が除去されているpUC18のEcoRI部位に、cDNA試料を連結した。イイ・コリ(E.coli)DH5αコンピテント・セル(ベセスダ・リサーチ研究所(Bethesda Research Laboratories))を形質転換し、TSWVのcDNAインサートを含むクローンを、50μg/mlのアンピシリン、IPTGおよびX-galを含有する寒天培地上に平板培養することによって一次選抜した。アルカリ溶解法を用いて、選抜クローンからプラスミドDNAを単離し[ビイアールエル・フォーカス(BRL Focus)第11巻:7頁(1989年)参照]、EcoRI制限酵素消化に続くジーンスクリーン・プラス(GeneScreen Plus)・ナイロンフィルター(デュポン社(DuPont))上へのDNAトランスファーによってインサートのサイズを測定した。TSWV-BL S RNAのcDNAインサートを含むプラスミドクローンは、以下に記載するごとく、TSWV-CPNH1 S RNAのヌクレオチド配列(GCAAGTTCTGCGAGTTTTGCCTGCT)に相補的な32P-標識オリゴマー(AGCAGGCAAAACTCGCAGAACTTGC)に対するハイブリダイゼーションによって同定した[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:001頁(1990年)参照]。幾つかのクローンをアガロースゲル上で同定し分析して、インサートのサイズを測定した。クローンpTSWVS-23は、約1.7kbの長さの最も大きなcDNAインサートを含んでいることが判明した。
【0018】
ポリメラーゼ鎖反応(PCR)を使用することにより全長NP遺伝子を得た。第一鎖cDNA合成は、TSWV NP遺伝子の5'末端(TSWV-CPNH1のヌクレオチド2571ないし2773位)においてS RNAに相補的である(TSWV-10Wのヌクレオキャプシド遺伝子を合成するのにも用いた)オリゴマープライマーJLS90-46(5'->3')AGCTAACCATGGTTAAGCTCACTAAGGAAAGCを用いて、20μl反応混合液中、37℃にて30分間で行った。その反応混合物には、1.5μlのウイルスRNA、1μgのオリゴマープライマー、0.2mMの各dNTP、1×PCR緩衝液(ジーンアンプ・キット(GeneAmp kit)、パーキン-エルマー-シータス社(Perkin-Elmer-Cetus))、リボヌクレアーゼ・インヒビター中の20UのRNA(プロメガ社(Promega))、2.5mMのMgCl2、および25UのAMV逆転写酵素(プロメガ・コーポレイション(Promega Corporation))が含まれていた。95℃にて5分間加熱することによって反応を終了させ、氷上にて冷却した。次いで、各1μgのオリゴマープライマーJLS90-46およびJLS90-47(5'->3')、(TSWV-10Wのヌクレオチド遺伝子を合成するのにも用いた)(後者のオリゴマーは該遺伝子の3'非コーディング領域(TSWV-CPNH1の1919ないし1938のヌクレオチド位)のS RNAと同一である)を用いる業者の取り扱い説明書(パーキン-エルマー-シータス社(Parkin-Elmer-Cetus))に従って、10μlのcDNA/RNAハイブリッドを用いてNP遺伝子をPCR増幅させた。典型的なPCRサイクルは、92℃にて1分間(変性)、50℃にて1分間(アニーリング)、および72℃にて2分間(重合)であった。試料は、1.2%アガロースゲル上に直接負荷して分離させた。分離したNP遺伝子断片をアガロースゲルから抽出し、エタノール沈殿し、20μlの蒸留水に溶解した。
【0019】
実施例IV
植物発現ベクターおよび植物形質転換ベクターの構築
実施例IIIからゲル単離したNP遺伝子断片を、50μl反応緩衝液[50mMのトリス-HCl(pH8.0)、10mMのMgCl2、0.1MのNaCl]中の制限酵素NcoIで37℃にて3時間消化し、NcoIで消化した植物発現ベクターpB1525に直接クローン化した。得られたプラスミドを同定し、カリフラワー・モザイク・ウイルス(CaMV)35Sプロモーターに対してセンス向きであるプラスミドをpB1525-NP+と命名し、逆向きであるものをpB1525-NP−と命名した。パン(Pang)らにより記載されているごとく[ジーン(Gene)、第112巻:229頁(1992年)参照]、ニコチアナ・タバカム(Nicotiana tobacum)プロトプラストにおけるNP遺伝子の経時的発現により、この発現カセットのNP産生能を測定した。次いで、(NP遺伝子は内部HindIIIおよびEcoRI部位を含んでいるため)HindIII/EcoRIでの部分消化によって、NP遺伝子を含有する発現カセットをpB1525-NP+から切り出し、同酵素で切断した植物形質転換ベクターpBIN19(クロンテック・ラボラトリーズ,インコーポレイティド(Clontech Laboratories,Inc.))に連結した。ホルスターズ(Holsters)らにより記載されている方法[モレキュラー・アンド・ジェネラル・ジェネティクス(Mol.Gen.Genet.)第163巻:181頁(1978年)参照]を用いて、得られたベクターpBIN19-NP+および対照プラスミドpBIN19をエイ・チュメファシエンス(A.tumefaciens)LBA4404株に移入した。
【0020】
クローンpTSWV-23およびPb1525-NP+中のインサートのヌクレオチド配列分析は、ジデオキシ法、T7ポリメラーゼ(ユウエス・バイオケミカルズ社(U.S.Biochemicals)、シークエナーゼ(Sequenase)TM)、ならびにシーミニアック(Siemieniak)らにより記載されている二重鎖配列決定法[アナリティカル・バイオケミストリー(Analyt.Biochem.)第192巻:441頁(1991年)参照]を用いて測定した。ヌクレオチド配列は双方のDNA鎖から決定し、この情報をジェネティクス・コンピューター・グループ(ジイシイジイ(GCG),ワイオミング州・マジソン(Madison,WI))から販売されているコンピューター・プログラムを用いて、TSWV単離株CPNH1の公表されている配列と比較した。
【0021】
タバコ・プロトプラストにおけるNP遺伝子の経時的発現も調製した。大規模アルカリ法(large scale alkaline method)を用いて、クローンpTSWVS-23および(PCR-構築NP遺伝子インサートを含んでいる)pUC18cpphasTSWV-NPについてのプラスミドDNAを単離した。NcoI消化によってPCR-構築NP遺伝子インサートをクローンpBIN25-NP+から切り出して、販売されている配列決定用のフランギング・オリゴマー・プライマーを利用した。発現カセットpUC18cpphasは、ファゼオラス・ブルガリス(Phaseolus vulgaris)種子貯蔵遺伝子であるファゼオリン由来のポリ(A)付加シグナルをそれが利用する点を除いて、pUC18cpexpに同様である。これらのプラスミドDNAを、ベックマンTi 70.1固定角ローターを用いて、2つのCsCl-エチジウムブロミド勾配分画に付した。DNA配列は、前記したジデオキシ法および二重鎖プラスミドDNA配列決定法を用いて得た。ヌクレオチド配列反応物を1m長のサーモスタット制御された(55℃)配列決定用ゲル上で電気泳動し、平均約750bpのヌクレオチド配列の読みを得た。ヌクレオチド配列は、双方のクローン化インサートの双方のDNA鎖から決定して、精度を確認した。TSWV-BL S RNA単離株からのヌクレオチド配列情報を、以下に論じるごとく、コンピューター・プログラム(ジイシイジイ(GCG)、ワイオミング州・マジソン(Madison,WI))を用いてTSWV単離株CPNH1およびL3と比較した。
【0022】
TSWV-BL S RNAのクローン化cDNAおよびPCR-構築インサートのヌクレオチド配列および予想アミノ酸配列ならびに、それらとTSWV-CPHN1 S RNAのヌクレオチド配列との比較を以下に示す。シーミニアック(Siemieniak)の二重鎖ジデオキシ配列決定法を用いてTSWV-BL S RNAクローンpTSWVS-23(TSWV-23)およびpBI1525-NP+(TSWV-PCR)のヌクレオチド配列を得、ジーンバンク(GeneBank)登録番号D00645に報告されているTSWV-CPNH1 S RNAのヌクレオチド配列の関連領域と比較した。TSWV-CPNH1 S RNAのヌクレオチド配列は、ド・ハーン(De Haan)(1990年)により報告されており、以下の配列によって表される:
【0023】
【化1】

【0024】
TSWV-CPNH1 S RNA上の非構造蛋白質遺伝子の不完全な予想アミノ酸配列は、1位の核酸で始まり783位で終了する核酸コドンで終了し、これを以下に掲げる:
【0025】
【化2】
【0026】
以下に図示するTSWV-23のヌクレオチド配列は、前記で得られたTWSV配列と酷似しており、非構造遺伝子の1/2とヌクレオキャプシド蛋白質遺伝子の1/2を含んでいる。
【0027】
【化3】
【0028】
以下に図示する本発明によるTSWV-PCRの核酸配列も、前記で得られたTSWV配列と酷似しており、ヌクレオキャプシド蛋白質遺伝子の全体をカバーしている。
【0029】
【化4】
【0030】
合わせて、クローン化TSWV-23インサートはTSWV-PCRインサートと重複しており、合わせて、それらは本発明によるTSWV-BL S RNAの2028個のヌクレオチドを表している。本発明によるこの2028個のヌクレオチド配列は、非構造遺伝子の一部分とヌクレオキャプシド蛋白質遺伝子の全体を含んでいる。合わせた配列は以下の通りである:
【0031】
【化5】
【0032】
この比較により、クローンpTSWVS-23のcDNAインサートが約760bpの52K蛋白質のウイルス成分遺伝子、完全な遺伝子間(intergenic)領域(492bp)、および450bpのNP遺伝子(NP遺伝子の約半分)を含んでいることが示された。このクローン化インサートは、EcoRI認識部位に正確に位置するその3'-末端を有しており、このことはcDNAクローニング工程の間の不完全なEcoRIメチル化を示している。このクローンは完全なTSWV-BLのNP遺伝子を含んでいないにも拘わらず、PCR遺伝子構築NP遺伝子の配列と450bPの重複をそれが有するため(TSWVにつき、全2028bpのTSWV-BL S RNAがヌクレオチド配列中に示されている)、その配列はかなり重要であった。このTSWV-BL PCR-構築およびTSWV-CPNH1 NP 遺伝子の間の配列比較により、合計21ヌクレオチドの相違(2.7%)、およびその内の8個がアミノ酸置換(3.1%)をコードしているが明らかとなった。突然変異が導入されることが知られているTaqポリメラーゼを用いてこのPCR構築NP遺伝子を得たため、これらの相違のうち幾つかはPCR増幅の間に導入された可能性がある。しかしながら、これらのヌクレオチド相違のうち15個は、TSWV-BL cDNAおよびPCRクローンの間の重複領域内に位置し、これらのヌクレオチド相違の1つ(TSWVの1702位;TSWV-PCRの485位)を除く全ては、双方のTSWV-BL S RNA起源のクローンによって共に保有されている。この比較は、これらの2つのクローン化NP遺伝子領域のヌクレオチド配列間の差異に全くそうではないにせよ大いには寄与しないことを明らかに示した。1702位におけるヌクレオチドの差異の結果、IleとSerとのアミノ酸置換が起こり、この差異でさえTSWV-BL単離株内の相同性の欠如によるものであろう。
【0033】
実施例V
アグロバクテリウム媒介形質転換:
アグロバクテリウム株LAB4404(クロン・テク(Clon Tech))の液状培地中に一晩浸漬することによって、ベクターpBlN19−NP+または対照プラスミドpBlN19を含有する該アグロバクテリウム株と共にニコチアーナ・タバカム・バール・ハバナ・シイブイ(Nicotiana tabacum var Havana cv)423の葉ディスクをインキュベートし、該接種したリーフディスクを非選択的MS培地上で3日間インキュベートした[サイエンス(Science)第227巻:1229頁(1985年)参照]。形質転換した細胞を選択し、苗条再生のために、300μg/mlカナマイシンおよび500μg/mlカルベニシリンを含有するMS培地中で再生させた。小植物を無ホルモン培地に移した後、根を誘導した。根が出た形質転換体を土壌に移し、温室条件下で成長させた。該MS培地は、十分な強度のMS塩(シグマ社(Sigma))、30g/lのグルコース、1mg/lのBAおよび1mlのB5ビタミン[1mg/mlのニコチン酸、10mg/mlのチアミン(HCl)、1mg/mlのピリドキシン(HCl)、100mg/mlのMyo−イノシトール]を含有する。トランスジェニック植物を自家受粉させ、種子をカナマイシン培地上で選択的に発芽させた。
【0034】
実施例VI
蛋白質の血清学的検出:
二重抗体サンドイッチ酵素結合免疫吸着法(DAS−ELISA)を用いて、TSWV−BL NPに対するポリクローナル抗体でトランスジェニック植物におけるNP遺伝子の発現を検出した。各試料は、酵素結合緩衝液[リン酸緩衝セーライン、0.05%ツイーン(Tween)20、2%ポリビニルピロリドン40、および0.2%オバルミン]の3ml中で、植物の頂部第2葉からのリーフディスク(約0.05g)を摩砕することによって調製した。タバコ・プロトプラストについては、遠心後の細胞抽出物をアッセイで直接用いた。トランスジェニック植物およびタバコ・プロトプラスト双方からの試料の10倍および3倍希釈物をDAS−ELISAの直前に作成した。
【0035】
ウェスタンブロットについては、2×SDS/試料緩衝液(0.126M トリス緩衝液、20%グリセロール、2%SDS、2%2−メルカプトエタノール、および0.01mg/mlブロモフェノールブルー)の0.25ml中でリーフディスク(約0.05g)を摩砕した。該ホモジネートを遠心し、負荷の前に上清を沸騰させた。蛋白質(10〜20μl試料/レーン)を分離し、膜上にブロットした。次いで、製造業者のimmunoselectキット指示マニュアル(ジブコ・ビイアールエル・ライフ・テクノロジーズ・インコーポレイテッド(Gibco BRL Life Technologies Inc.))に従って該膜を加工した。全ウイルス粒子に対する抗体を健康なタバコ植物からの細胞抽出物で予め吸収させ[プラント・ディジーズ(Plant Disease)第70巻:501頁(1986年)参照]、2μg/mlの濃度にてウェスタンブロットで用いた。
【0036】
TSWV−BLウイルス粒子、またはTSWV−BLもしくはTSWV−1に対して生起させた抗体を用い、TSWV単離株(TSWV−BL、アルカンサス、10W体菜、ベゴニアまたはブラジル)の血清学的反応をDAS−ELISAで検定した。
【0037】
実施例VII
TSWV単離株でのトランスジェニック植物の接種
接種物は、ニコチアナ・ベンタミアナ・ドミン(Nicotiana benthamiana Domin.)を異なるTSWV単離株で感染させ、緩衝液(0.003M KH2PO4、0.067M K2HPO4および0.01M Na2SO3)15ml中でエヌ・ベンタミアナ植物の感染させた葉(0.5g)を摩砕する(接種1ないし2週間後)ことによって調製した。接種抽出物を、トランスジェニック植物のコランダム散布葉上で直ちに擦り、接種した葉を引き続いて水で濯いだ。TSWVは摩砕後イン・ビトロでは高度に不安定であるので、接種物の各バッチを用いて、NP遺伝子を含有するNP(+)植物をまず接種し;各接種物の最後の接種植物は、常に、ベクター配列単独を含有する対照NP(-)植物であって、特定のウイルス接種物はなお接種の最後において感染性であることを確実とした。
【0038】
接種7〜15日後に、局所病巣および全身感染についてのデータを採り、特に示す以外は、接種した植物の数にわたる全身感染した植物の数として以下の表に表した。この表において、「ELISA」の下で収集したデータは、それからR1植物が由来するR0系のデータであり;ベゴニア単離株はR1植物に局所病巣を誘導し、および抵抗性は、接種した植物の数にわたる局所病巣生成植物の数として表し;NTはテストをしなかったことを示す。
【0039】
ウイルス(TSWV)をスポットしたタバコのヌクレオキャプシド蛋白質
(NP)遺伝子を発現するR1植物のTSWV単離株での接種に対する反応
TSWV単離株に対する反応
エライザ
(R0pl.) BL アルカンサス 10W体菜 ベゴニア ブラジル
R0系
NP(+) 0.015 0/10 4/25 3/24 29/40 36/36
NP(+)4 0.386 6/30 21/23 18/21 9/48 42/42
NP(+)9 0.327 0/20 NT 20/20 - -
NP(+)14 0.040 0/20 - 9/20 8/18 18/18
NP(+)21 0.042 0/15 5/15 3/15 2/4 6/6
NP(+)22 0.142 0/20 - 15/20 31/36 36/36
NP(+)23 0.317 0/20 - 16/20 - -
NP(-) - 42/42 24/24 62/62 66/66 54/54
【0040】
前記したごとく、TSWVのS RNA成分中に存在するTSWV−BL NP遺伝子の単離株は、2つの戦略を用いてアプローチした。cDNAクローニング戦略は、TSWV−CPNH1 S RNAに相補的なオリゴマープローブに対するハイブリダイゼーションによって同定したところによると、TSWV−BL S RNAに由来するcDNAインサートを含有するいくつかのクローンを生じた。クローンpTSWVS−23は、約1.7kb長さにマップされる最長インサートを含有していた。第2の戦略は、TSWV−CPNHl S RNAの公表された配列、および直接全TSWV−BL RNAからの発現のためにNP遺伝子を増幅し、作成するPCRを利用した。オリゴマープライマーJLS90−46および−47を合成し、JLS90−46はNP遺伝子の5’暗号領域中のS RNAに相補的であり(TSWV−CPNH1の2051〜2073位)、一方、JLS90−47はNP遺伝子の3’非暗号領域のものである(TSWV-CPNH1の1218〜1237位)。両プライマーは、引き続いてのクローニングのための制限酵素Nco1についての認識部位を含有し、プライマーJLS90−46は植物コンセンサス翻訳開始コドン配列(AAXXATGG)を有し、これは、翻訳に際して、NP遺伝子の第3コドン(GTT)に翻訳開始コドンを融合させることが期待された。翻訳開始コドンのTSWV−BL NP遺伝子の第3コドンへの融合を行って、いずれのアミノ酸コドンも取り込むことなくNcoI認識部位を保持した。かくして、PCTで作成したTSWV NP遺伝子の発現は、天然NPよりもN末端で2のアミノ酸(Ser-Lys)だけ短いTSWV-BL NPを生じるであろう。
【0041】
この特異的に増幅させた約850bpのDNA断片をNcoIで消化し、植物発現ベクターpB152Sにクローンした。CaMV 35Sプロモーターに対するTSWV−BL NP遺伝子の向きは、制限酵素部位マッピング(EcoRI、HindIII、AvaIおよびAiwN1)によって決定した。適当な向きでインサートを含有するいくつかのクローンを単離し(pB1525−NP+)、他のものは反対の向きでインサートを含有した(pB1525−NP−)。また、この制限酵素部位マッピングは、クローンpB1525−NP+のインサートが、TSWV-CPNH1 NP遺伝子で見い出されるものと同一の制限酵素部位を含有することを示した。かくして、TSWV−BL NP遺伝子の発現は、発現ベクターpB1525のアルファルファモザイクウイルス(ALMV)の5’−非翻訳リーダー配列に融合した二重CaMV 35Sプロモーターによって制御された。重ねた二重CaMV 35Dプロモーター要素を利用する発現ベクターは、単一の35Sプロモーター要素を利用する同様のベクターよりも高レベルのmRNA転写を生じる。
【0042】
3つのpB1525−NP+クローンを一時的にタバコ・プロトプラストで発現させて、増幅されたDNA断片がNPをコードすることを確認した。これを達成するために、PEG法によってクローンをタバコ・プロトプラストに移入し、インキュベーションの2日後、全TSWV−BLウイルス粒子に対する抗体を用い、DAS−ELISAによって発現されたNPを検出した。プラスミドpB1525-NP+中にNP遺伝子を保有するタバコ・プロトプラストで高レベルのNPが産生され:一方、アンチセンスNP配列で形質転換したタバコ・プロトプラスト(pB1525−NP−)ではNPは検出されなかった。
【0043】
前記したごとく、クローンpB1525−NP+のPCR構築インサートおよびクローンpTSWV−23のtehcDNAインサートを二重鎖配列決定に付した。cDNAおよびPCRクローンの配列解析により、各々、1.71kbおよび865bpのインサートが明らかにされ、これは、配列TSWV−CPNH1 S RNAと比較した場合、クローンpTSWV−23のcDNAインサートが52K蛋白質ウイルス粒子成分遺伝子、完全な遺伝子間領域(492bp)およびNP遺伝子の450bp(該遺伝子の約半分)を包含することを示す。このクローン化インサートはEcoRI認識部位に正確に位置するその3’末端を有し、これは、cDNAクローニング間における不完全なEcoRIメチル化を示唆する。このクローンは完全なTSWV−BL NP遺伝子を含有しないにも拘わらず、その配列は非常に重要である。というのは、それはPCR構築NP遺伝子の配列と450bpの重複を有するからである。このTSWV−BL PCR構築NP遺伝子とTSWV−CPNH1 NP遺伝子の間の配列の比較は、合計21個のヌクレオチドの差異(2.7%)を明らかにし、そのうち8つはアミノ酸置換をコードする(3.1%)。このPCR−構築NP遺伝子は、突然変異を取り込むことが知られている、Taqポリメラーゼを用いて得られたので、これらの差異のうちいくらかはPCR増幅の間に導入することができる。しかしながら、これらのヌクレオチド差異のうち15はTSWV−BL cDNAクローンおよびPCRクローンの間の重複領域内に位置し、これらの差異のうちの1つ(1702位)を除いたすべてが、両TSWV−BL R RNA由来クローンに存在する。この比較は、明らかに、PCR増幅は、これらの2つのNP遺伝子のヌクレオチド配列間の差異に全てではないにせよ大いには寄与しないことを示す。1702位におけるヌクレオチドの差異の結果、IleのSerでのアミノ酸置換が起こり、この差異でさえTSWV−BL単離株内における相同性の欠如によるであろう。
【0044】
ヌクレオチドの差異がTSWV単離株間の発散に帰されるという可能性は、
TSWV−CPNH1、TSWV−L3、およびTSWV−B1 S RNAの間の他の配列決定された領域との比較によって支持される。これらの比較は以下の表に示す:
【0045】
【表1】
【0046】
CPNH1およびL3単離株からのNP遺伝子のヌクレオチド配列は相互に3.1%だけ相違し、BL単離株とは、ほとんど同程度(2.5%)相違する。しかしながら、CPNH1およびBL単離株の間のNPアミノ酸配列は、L3およびBLの間、またはCPNH1およびL3単離株の間よりも、かなり相違する。また、前記表の結果は、これらのTSWV単離株のNP遺伝子領域が、52K蛋白質のアミノ酸配列間の差異が7.9ないし10.6%の範囲であるので52K蛋白質よりも高い程度の選択圧に付されており、NPのアミノ酸配列について見い出されているよりも2倍以上であることを明らかにする。ヌクレオチド配列の発散は遺伝子間領域の間で最大であり、これは、この領域が遺伝子領域よりも低い選択圧に付されてことを示す。
【0047】
トランスジェニック植物におけるNP遺伝子配列の存在は、まず、PCR分析によって確認された。NP遺伝子を挟む配列に相同なプライマーを用い、約800bpのNP DNA断片がトランスジェニックNP(+)植物の全DNAから特異的に増幅され、一方、対応する断片は対照NP(-)植物では検出されなかった。NP遺伝子の発現はDAS−ELISAによって各R0トランスジェニック植物でアッセイし、結果は以下の表に示す:
【0048】
トマト黄化壊疽ウイルス(TSWV)のヌクレオキャプシド蛋白質(NP)遺伝子を発現するR0トランスジェニック植物のTSWV-BL単離株での接種に対する反応
植物年齢 R0クローン ELISAa 病巣/葉b NP(+):NP(-)c
7〜8葉 NP(+)1 0.374 7(199) 1:28
NP(+)2 0.015 0(199) 0:199
NP(+)3 0.407 23(102) 1:4
NP(+)4 0.386 2(102) 1:51
NP(+)5 0.023 0(124) 0:124
NP(+)6 0.197 35(325) 1:9
NP(+)7 0.124 1(325) 1:325
9〜10葉
NP(+)8 0.344 36(36) 1:1
NP(+)9 0.327 2(20) 1:10
NP(+)10 0.406 34(33) 1:1
NP(+)11 0.156 5(20) 1:4
NP(+)12 0.133 9(57) 1:6
NP(+)13 0.144 2(7) 1:4
NP(+)14 0.040 0(19) 0:19
NP(+)16 0.053 0(10) 0:10
5〜6葉
NP(+)20 0.487 203(117) 2:1
NP(+)21 0.042 0(117) 0:117
NP(+)22 0.142 0(208) 0:208
NP(+)23 0.317 223(208) 1:1
NP(+)24 0.051 0(35) 0:35
NP(+)25 0.286 13(35) 1:3
NP(+)26 0.037 0(22) 0:22
NP(+)27 0.425 305(22) 14:1
a トランスジェニック植物におけるNPの産生は二重抗体サンドイッチ酵素結合免疫吸着法(DAS−ELISA)によって検定した;コーティング用のウイルス粒子に対する抗体の濃度:1μg/ml;TSWV−BLへのコンジュゲートの希釈:1:250;基質添加後150分に採取した結果;405nmにおける読み
b 接種した葉で発達した局所病巣は接種7日後に計数した。データは3つの接種した葉の平均を表す。括弧に入れたデータは、同一接種物で接種した対照NP(+)植物から生じた病巣の数である。
c pBIN19−NP+
で形質転換したNP(+)植物で発達した局所病巣-対-同一接種物で接種した場合の対照NP(-)植物で発達した局所病巣の比率
【0049】
23のNP(+)クローンのうち、10は高レベルのNPを産生し、5は中レベルのNPを蓄積し、および残りの8は低レベルのNPを産生した。トランスジェニック植物で発現されたNPのサイズはウェスタンブロット法によって分析した。タバコ抽出物からの多くのポリペプチドは、抗体を健康なタバコ植物からの抽出物で予め吸収させたにも拘わらず、全ウイルス粒子に対する抗体に対して反応性であった。それらのうち、1つのバンドのみが、NP遺伝子で形質転換したタバコ植物からのポリペプチドのパターンに対してユニークであった。このポリペプチドは約29kDaであると評価され、これは、天然NPの予測されるサイズに近い。同様のサイズの抗体反応性蛋白質バンドは、ベクターpBIN19を含有するトランスジェニック植物からの抽出物では見い出されなかった。
【0050】
TSWV−BL単離株でのタバコ葉の接種の結果、天候条件および植物の生理学的段階に応じて、全身感染または壊死性局所病巣いずれかとなるであろう。R0植物をウイルス抵抗性につきTSWV−BLでテストした場合、TSWV−BLは、接種6〜8日後に、対照NP(-)植物の接種した葉上に典型的な壊死性病巣を誘導した。しかしながら、トランスジェニックNP(+)植物は、対照NP(-)植物に比して、ウイルスに対する抵抗性スペクトルを示した。23のNP(+)植物のうち7つはいずれの局所病巣も発生せず、あるいは発生した病巣の数は対応する接種したNP(-)植物上のものよりも少なくとも20倍小さかった。3つのNP(+)植物は中程度の反応を有していたが(対照よりも5−ないし19−倍小さい)、残りの9の植物は低い抵抗性であるか、または全く抵抗性がなかった。接種したNP(+)またはNP(−)植物のいずれも全身感染を示さなかった。無病徴R0植物をそのライフサイクルの最後までモニターし、そのライフサイクルを通じて病徴が観察されなかった。無病徴NP(+)植物の接種した葉を、シイ・キノラ(C. quinola)植物の葉上のウイルスの存在についてチェックした。高度に抵抗性のNP(+)植物のTSWV−BL−攻撃葉からウイルスは回収されず、これは、該ウイルスがこれらのNP(+)植物では複製もしくは増殖できないことを示す。
【0051】
選択したR0植物からのリーフディスクを継代クローンし、再生した小植物を該ウイルスによって攻撃した。すべての継代クローンしたR0植物は、それらの対応するオリジナルのR0植物と同様のレベルの抵抗性を示した。
【0052】
TSWVは広く分布して、多くの生物学的距離の株が存在するので、異なるTSWV単離株による感染に抵抗性を示すトランスジェニック植物の効果もテストした。5つのTSWV単離株を本実験で選択して、カナマイシン含有培地上で発芽したR1植物を攻撃した:TSWV−BL、アルカンサス、10W体菜、ベゴニアおよびブラジル。最初の3つの単離株は全ウイルス粒子およびTSWV-BL(通常のTSWV「L」血清群)に対する抗体に対して反応性であった(図5参照)。ベゴニア単離株はTSWV−IのNP(「I」血清群)に対する抗体と強く反応したが、TSWV−LP NPに対して生起したものに対しては反応せず、従って、「I」血清群に属した。ブラジル単離株の、TSWV−BLまたはTSWV−I血清群のNPに対する抗体に対する検出可能な反応は見い出されず、それは、TSWV−BLの全ウイルス粒子に対する抗体には弱い反応性であった。さらに、この単離株は感染したタバコおよびエヌ・ベンサミアーナ(N.benthamiana)の葉上で全身の斑および皺を引き起こしたが、カボチャまたはキュウリを感染せず、これは、それがウリ科植物単離株とは区別される単離株であることを示す。これらの結果は、この単離株が第3の血清群であると考えられることを示す。
【0053】
7つのR0系に由来する実生をカナマイシン培地上で発芽させ、前記したTSWV単離株で接種した。感染性データは接種7日後に開始し、毎日記録した。TSWV−BL、アルカンサス、10W体菜またはブラジル単離株で接種した植物は、もしウイルス病徴が未接種葉で観察されれば、感受性とした。ベゴニア単離株で接種した植物は、局所病巣が接種葉で観察されれば、感受性とした。というのは、この単離株はタバコで全身性感染を引き起こさないからである。すべての接種した対照NP(-)R1植物は、これらの5つの単離株による感染に対して感受性であった。それらは、ベゴニアで接種したトランスジェニックR1植物が接種葉で局所病巣を生じたに過ぎないことを除き、接種12日後に全身感染した。しかしながら、ほとんどすべてのNP(+)R1植物は、同種単離株TSWV-BLに対して高い抵抗性を示したが、かなり低いパーセントのNP(+)R1植物が異種単離株アルカンサス、10W体菜およびベゴニアに対して抵抗性であった。他方、系NP(+)4からのNP−高発現NP(+)R1植物のいくつかでは、病徴性発現においてわずかな遅延(1ないし2日)が観察されたにも拘わらず、7つのトランスジェニック系からのすべてのNP(+)R1植物はブラジル単離株に対して感受性であった。
【0054】
抵抗性R1植物は、そのライフサイクルを通じて無病徴のままであった。17のより低い病徴性のNP(+)植物の接種した葉を、チェノポジウム・キノア(Chenopodium quinoa)植物の葉上の逆接種によってウイルスの存在についてチェックした。無病徴NP(+)植物の接種した葉からウイルスは回収されず、これは、該ウイルスがこれらのNP(+)植物で複製もしくは増殖できないであろうことを示す。
【0055】
トランスジェニック植物におけるNPの蓄積レベルと異種TSWV単離株に対する抵抗性の程度との間の関係も調べた。
【0056】
前記したデータの分析は、低レベルNPのR0系に由来するR1植物は「L」血清群(アルカンサスおよび10W体菜)の異種単離株に最良の抵抗性を付与し、一方、高レベルNPのR0系からのR1は「I」血清群に属するベゴニア単離株に抵抗性であることを示す。例えば、低NP発現系NP(+)2、14および21からの接種したR1植物の平均76%はアルカンサスおよび10W体菜による感染に対して抵抗性であり、一方、これらの単離株に対する抵抗性は、高NP発現系NP(+)4、9および23からの同様に接種した植物の11%のみで観察された。他方、ベゴニア単離株は、低NP発現系NP(+)2、14および21からのR1植物の79%に感染したが、高NP発現系NP(+)4からのR1植物では19%のみに感染した。
【0057】
従って、低レベルのNP遺伝子を発現するトランスジェニックR1植物は単離株10W体菜(「L」血清群)での感染に対して高度に抵抗性であるが、ベゴニア単離株(「I」血清群)での感染に対しては抵抗性でなかったと結論された。対照的に、高NP発現R1植物はベゴニア単離株による感染に対して非常に抵抗性であったが、10W体菜からの単離株による感染に対しては抵抗性でなかった。
【0058】
かくして、異種単離株に対して抵抗性と個々の植物におけるNP発現との関係を正確に定量するのは興味深かった。本明細書中で報告する多数の接種実験において、トランスジェニック植物の葉試料はアルカンサスおよび10W体菜単離株での接種前に採取した。また、NP発現レベルと抵抗性との間の明らかな関係の観察を行った後に、ベゴニア単離株で接種した植物の非接種葉から試料を採取した。サンプリングの後者の方法は、ベゴニア単離株による感染からの干渉なくして行うことができた。何故ならば、この単離株はタバコにおいて全身感染を引き起こさないのみならず、TSWV−BL NPに対する抗体とも反応しないからである。単離したTSWV−BLのNPに対して生起した抗体を用い、DAS−ELISAによって、すべての試料を相対的NPレベルにつき検定した。図5および6はトランスジェニックR1植物(それらが由来するR0系を問わない)におけるNPレベルとアルカンサスおよび10W体菜単離株に対する、またはベゴニア単離株に対するそれらの抵抗性との間の関係を示す。非常に低いまたは検出不可能なELISA反応性(0〜0.05 OD405nm)を有するほとんどすべてのトランスジェニックR1植物はアルカンサスおよび10W体菜単離株(「L」血清群)による感染に対して抵抗性であるが、ベゴニア単離株(「l」血清群)に対しては感受性であった。対照的に、高ELISA反応性(0.4〜1.0 OD405nm)を与えるほとんどすべてのR1植物はベゴニア単離株に対して抵抗性であったが、アルカンサスおよび10W体菜単離株に対して感受性であった。
【0059】
ブドウ葉巻ウイルスで感染させた組織からのdsRNAの単離で首尾よく用いられたことがある方法[アクタ・ホルティカルチャー(Acta Horticulture)第186巻:51頁(1986年)、およびカン・プラント・ディス・スルブ(Can. Plant Dis. Surv.)第68巻:93頁(1988年)]の組合せを用い、TSWV−Bで感染させたエヌ・ベンサミアーナ(N.benthamiana)植物から二本鎖(ds)RNAを単離した。該dsRNAをcDNA合成につき選択した。というのは、この単離株からのウイルス粒子の単離は可能ではなかったからである[プラント・ディジーズ(Plant Disease)第74巻:154頁(1990年)参照]。TSWV-BのS RNAに特異的なcDNAライブラリーを作成するために、二本鎖S RNAをゲル精製し、メチル水銀処理によって変性し、ランダムプライマーを用いてプロメガ社によって提供されたcDNA合成手法に付した。合成したcDNA断片をEcoRIアダプターを介してEcoRI消化のλ ZAPII(ストラタジーン社(Stratagene)) にクローン化し、ゲル精製S RNAの逆転写によって調製したcDNAプローブを用いるコロニーハイブリダイゼーションによって陽性クローンを同定した。数ダースの陽性クローンをアガロースゲル上で分析し、ほとんど全部のTSWV−B S RNAをカバーする、最長インサート(L1、L22およびL30)を含有する3つの重複クローンのみを選択した(図3参照)。
【0060】
クローンL1、L22およびL30中のインサートのヌクレオチド配列は、双方のDNA鎖から、最初はユニバーサルおよび逆プライマーによって、次いでTSWV−BのS RNAの配列決定のために設計された内部プライマーによって決定した。配列決定はサンガー(Sanger)のジデオキシ法、T7ポリメラーゼ(ユウエス・バイオケミカルズ(U.S.Biochemicals)、シークエナーゼ(Sequenase)TM)、およびシーミニアック(Siemieniak)によって記載された二本鎖の配列を決定する方法(アナリティカル・バイオケミストリー(Analyt.Biochem.)参照)を用いて行った。これらのクローンの配列解析は、それぞれ、1.994kb、2.368kbおよび1.576kbのインサートを示し、これらの配列はS RNAゲノムの93%を示した(図3参照)。組み立てた配列をジェネティクス・コンピューター・グループ(Genetics Computer Group)(ジイシイジイ、ウィスコンシン州(GCG,Madison,WI))から販売されているコンピューター・プログラムを用いてTSWV単離株CONH1、L3、IおよびBLの配列と比較することにより解析した。
【0061】
コンピューター分析によって、2.842kbの組み立てた配列は完全な52K非構造蛋白質遺伝子、完全な遺伝子間領域(629bp)、および737bpのNP遺伝子(該NP遺伝子の39のN−末端タクレオチドのみが表されなかった)をカバーすることが示された。N遺伝子のこの失われた領域を得るために、N遺伝子の開始コドンからの62個のヌクレオチド配列と同一のプライマーTTCTGGTCTTCTTCAAACTCAをポリヌクレオチドキナーゼで末端標識して前記したcDNAライブラリーをスクリーニングした。5つの推定クローンが得られた。5つのクローンの配列解析は、クローンS6およびS7のみがN遺伝子のこれらの39の失われたヌクレオチドを含有することを示した。また、後者のクローンはS RNAの3’極末端を包含した。
【0062】
S RNAの5’極末端は、5’RACEシステム(ジブコ(GIBCO))を用いて得られた。TSWV−BのssRNAおよびTSWV−Bで感染させたタバコ植物から単離した全RNAを共に用いて、te TSWV−B S RNAのヌクレオチド746−763位に相補的なオリゴヌクレオチド(5'-CTGTAGCCATGAGCAAAG)を持つ第1鎖cDNAを合成した。ターミナルデオキシヌクレオチジルトランスフェラーゼを用いて第1鎖の3'−末端にdCTPをテールした。次いで、ホモポリマーテールにアニールするアンカープライマー、およびTSWV−B S RNAのヌクレオチド512−529位にアニールするオリゴヌクレオチド(5'-TTATATCTTCTTCTTGGA)を用い、PCRによって、テールしたcDNAを増幅した。該PCR増幅断片をゲル精製し、配列解析のために、T−ベクターpT7Blue(ノバゲン(Novagen))に直接クローン化した。8つの独立したクローンを、S RNAの5’領域(TSWV−B S RNAのヌクレオチド40−57位)に密接なオリゴマープライマー(5'-GTTCTGAGATTTGCTAGT)で配列決定した。得られたクローンのうち6つはS RNAの5’極末端を含有し、これらのクローンからの5'-末端ヌクレオチド配列は同一であった。かくして、TSWV−B S RNAの完全なヌクレオチド配列は3049ヌクレオチド長であった。
【0063】
かくして、3つの従前に配列決定したクローンと共にこれらの2つのクローン(L1、L22、L30、S6およびS7)は、前記した全3032ヌクレオチドをカバーしていた。TSWV−CPNH1およびTSWV−1の末端配列との比較により、18ヌクレオチドの5'極末端は組み立てられた配列中には表されていないにも拘わらず、TSWV−B S RNAの3’−極末端はTSWV-I S RNAの3'-極末端と同一であり、TSWV−CPNH1の3'−極末端とは15ヌクレオチド異なることが明らかとなった。TSWV単離株間の末端配列の保存は、ブニヤウイルス属の他のメンバーの保存し一致し、これは、末端配列は安定な塩基対合構造を形成し、それはその複製およびキャプシド化に関与し得るという仮説を支持する。
【0064】
本発明によるTSWV−B(前記したブラジリアン単離株)のS RNAゲノムの完全なヌクレオチド配列は以下の通りである。
【0065】
【化6】
【0066】
【化7】
【0067】
本発明による非構造(前記単一下線)およびヌクレオキャプシド蛋白質の帰結されるアミノ酸配列は以下の通りである。
【0068】
【化8】
【0069】
および
【0070】
【化9】
【0071】
前記したヌクレオキャプシド蛋白質遺伝子はウイルスの相補鎖上にあり、TSWV−Bのヌクレオキャプシド蛋白質遺伝子は以下の通りである。
【0072】
【化10】
【0073】
TSWV−Bの完全なS RNAは3049ヌクレオチド長であるべきであり、TSWV−CPNH1のS RNAよりも134ヌクレオチド長い。この差異はTSWV−B S RNAの長い遺伝子間領域に主として帰せられる。TSWV−B S RNAの配列決定した領域の解析により、前記した2つのオープンリーディングフレームが明らかとされ、これは他のTSWV単離株と同様である。より大きい方は、ヌクレオチド88に始まり、ヌクレオチド1491で終わるウイルスRNA鎖上に位置していた。ウイルス相補鎖上のより小さい方は、ヌクレオチド2898における開始コドンおよびヌクレオチド2122の終止コドンによって境界が決せられた。2つのオープンリーディングフレームは629ヌクレオチドの遺伝子間領域によって分離されていた。TSWV−B S RNAの配置したヌクレオチドおよびアミノ酸配列と他の単離株のそれとのパーセント相同性比較を示すところの、全配列決定したTSWV−B S RNAと以下の表中の他の単離株のS RNA領域との比較は以下の通りである。
【0074】
全53K蛋白質遺伝子 遺伝子間 29K蛋白質遺伝子
比較a nt nt aa nt nt aa
B/CPNH1 76.4b 80.0 86.1(78.3)c 72.4 77.5 91.5(79.1)
B/L3 75.8 79.0 89.0(82.0) 76.4 78.0 91.1(79.9)
B/BL 76.3 - - 72.8 77.6 90.3(79.5)
B/I 63.0 - - - 63.1 69.7(55.3)
CPNH1/L3 94.8 95.6 92.0(89.4) 89.2 96.8 99.6(98.5)
CPNH1/BL 96.4 - - 95.9 97.2 98.8(96.9)
CPNH1/I 62.7 - - - 60.8 69.5(55.1)
L3/BL 95.1 - - 92.6 97.3 99.2(98.5)
L3/I 60.9 - - - 60.9 69.5(55.1)
I/BL 61.7 - - - 60.9 68.8(53.9)
a 単離株TSWV−CPNH1(2.916kb)、TSWV−L3(2.837kb)、TSWV−BL(2.037kb)およびTSWV−1(1.144kb)の部分的および完全なS RNA配列を、TSWV−BのS RNA配列(3.049kb)との比較のために用いた。
b パーセント類似性は、GCG配列決定分析ソフトウェアパッケージのプログラムBESTFITを用い、それらのヌクレオチドまたは予想されるアミノ酸配列の比較によって計算した。
c パーセント同一性は括弧に入れた。
【0075】
前記したごとく、最大のヌクレオチド配列の類似性(75.8%−76.4%)はL−タイプの単離株(CHNH1、L3およびBL)で示された。より程度が低いが、TSWV−B S RNA とI血清群に帰属させられたTSWV−IのS RNAの間にヌクレオチド配列類似性(63%)があった。比較すると、L-タイプの単離株(CHPN1、L3およびBL)の配列決定したS RNA領域は94.8%−96.4%のヌクレオチド配列類似性を保有していた。
【0076】
777ヌクレオチドのオープンリーディングフームは予測される分子量が28700Daの258アミノ酸のN蛋白質をコードする。TSWV単離株からのNオープンリーディングフレームの配列比較により、TSWV単離株CPNH1、L3およびBLからのN遺伝子のヌクレオチド配列は、それらが相互に異なる(2.7%−3.2%)よりも、TSWV−Bとはかなり(22%−22.5%)異なっていることが明らかにされた。免疫学的分析の結果と一致して、CPNH1、L3およびBL単離株のNアミノ酸配列は、TSWV−Bに対する(90.3%−91.5%の類似性または79.1%−79.9%の同一性)よりも、相互に対してより密接である(98.8%−99.6%の類似性または96.9%−98.5%の同一性)。かなり低い相同性が、ヌクレオチド(63.1%)およびアミノ酸(69.7%の類似性または55.3%の同一性)レベル双方においてTSWV−Iに対して観察された。262アミノ酸をコードするTSWV−IのNオープンリーディングフレームを除き、他の単離株のNオープンリーディングフームは258個のアミノ酸についてコードする。コンピューター解析は、TSWV−I Nオープンリーディングフームの余分な残基はアミノ酸配列の挿入の結果であることを示す(残基82ないし84および残基116)。1つの可能なN−グリコシル化部位が残基68に見い出される。
【0077】
1404ヌクレオチドの第2のオープンリーディングフレームは、分子量が52566Daと予測される467個のアミノ酸の非構造蛋白質をコードする。TSWV−CPNH1およびTSWV−L3の相同オープンリーディングフレームの比較は、ヌクレオチドレベルで80%および79%の類似性、ならびにアミノ酸レベルで86.1%(または78.3%の同一性)および89%(または82.0%の同一性)の類似性を示した。このオープンリーディングフレームは4つの可能なグリコシル化部位を含有し、これはTSWV-CPNH1およびTSWV-L3と正確に同一の位置に存在する。
【0078】
いくつかの挿入のため、TSWV−B S RNAの遺伝子間領域は、TSWV-CPNH1およびTSWV−L3の対応物よりも各々126および41ヌクレオチド長かった。プログラムFOLDによる配列解析は、遺伝子間領域は、当該遺伝子間領域のA−リッチのストレッチと共にU−リッチのストレッチを内部的に塩基対合させることによって、非常に複雑で安定なヘアピン構造を形成することができ、これは、最小自由エネルギー値によって表してTSWV−CPNH1およびTSWV−L3から得られたものと同様の安定性を有することを示した。この内部塩基対合構造は転写終始シグナルとして働き得る。
【0079】
また、前記表の結果は、TSWV−BのN蛋白質は52K蛋白質よりも高い選択圧に付されていることが明らかとなった;52K蛋白質のアミノ酸配列の間の類似性はNPのアミノ酸配列で見い出されるものよりも低い。ヌクレオチド配列の発散は遺伝子間領域のうちで最も高く、これは、この領域がいずれの遺伝子領域よりも高い選択圧に付されていることを示す。
【0080】
TSWV−Bおよび他の4つのTSWV単離株の間の発展的な関係を解析し、これを図4に示すが、そこでは、発展的な系統樹組織はこれらのTSWV単離株について収集された血清学的データの関連性と合致する。かくして、本発明におけるTSWV−Bは、I−タイプの単離株TSWV−Iに対するよりもL−タイプの単離株に対してより密接であるが、L−タイプの単離株が相互に対するよりもL−タイプの単離株に対して類似性がかなり低い。
【0081】
病徴性発現のわずかな遅延にも拘わらず、トランスジェニック植物はTSWVのブラジル単離株に対して抵抗性を示さなかった;血清学的結果は、この単離株が「L」および「I」タイプの単離株から区別され、ウリ科植物単離株から生物学的に異なることを示す。かくして、ブラジル単離株は、なお、TSWVのもう1つの血清群に属するであろう。いずれにせよ、感染性の結果は、単一のNP遺伝子がトスポウイルス属におけるすべての単離株に対する抵抗性を提供するようである。
【0082】
低いまたは検出できないELISA反応性(0−0.05 OD405nm)を与える本発明によるトランスジェニック植物は、「L」血清群の異種単離株(アルカンサスおよび10W体菜)による感染に対して抵抗性であり、一方、これらの単離株に対する保護は高レベルのNPを蓄積する植物では見い出されなかった。対照NP(−)植物のELISAの判読値(0.05 OD405nm)に比較して、これらのトランスジェニック植物はもしあってもほとんどTSWV−BL NPを生産しないであろう。同様の結果がトランスジェニック植物で観察されており、そこでは、CPの蓄積は検出されず:これらはウイルス感染に対して高い抵抗性を示した。この現象を基礎づける機構は現在知られていない。このタイプの抵抗性は、恐らくは、攻撃ウイルスのマイナスセンス複製RNAにハイブリダイズさせ、必須の宿主因子(例えば、レプリカーゼ)に結合するかあるいはウイルス粒子組立体に干渉することによって、トランスジェニック植物で生じるCP RNA分子とウイルス複製との干渉に帰すことができるようである。
【0083】
しかしながら、同種TSWV−BL単離株に対する抵抗性は、明らかに、NP遺伝子の発現レベルから独立していることに注意されたい。TSWV−BLで接種した個々のR1植物の相対的NPレベルは測定しなかったが、これらの接種R1植物(合計145のテスト植物)で生じたNPは検出不可能ないし高度の範囲であると仮定するのは合理的である。
【0084】
「L」血清群の異種単離株からの保護の場合とは対照的に、TSWV−I血清群のベゴニア単離株からの保護が高NP発現R1植物で見い出された。「L」血清群のNPヌクレオチド配列と「I」血清群のそれとの比較により、ヌクレオチドおよびアミノ酸における各々62%および67%の同一性が明らかにされた。2つの血清群のNP遺伝子の差異は余りにも大きくて、トランスジェニック植物で生じたNP(「L」血清群)は、「I」血清群のベゴニア単離株を攻撃するに際して機能不全蛋白質として働くであろう。この「欠陥」外皮蛋白質のウイルス粒子への取込みは、ウイルスの運動またはそのさらなる複製を阻害する欠陥ウイルスを生じるであろう。このタイプの相互作用は、保護には高レベルのNPを要すると予測される。また、ベゴニア単離株に対する抵抗性は、R1植物で産生されるNP転写体の、ウイルス複製での干渉を含む。もしこれが真実であれば、異種ウイルスの複製を阻害するのに、(2つのNP遺伝子の異種性質のため)より多くのNP転写体を要するであろう。
【0085】
「L」および「I」血清群の異種単離株に対する抵抗性に対する個々のR1植物におけるNPレベルの関係を示す結果について明らかな説明はないが、これらは明らかな傾向であると考えられる。というのは、データは多数(190)の植物から得られたものだからである。かくして、個々の植物におけるCPまたはNPレベルの測定はNPまたはCPレベルを抵抗性に関係付けるためのより正確な方法を提供すると考えられる。この形式のデータ解析によって、抵抗性は、それらが由来するR0系のNPレベルに対するよりも、各テスト植物におけるNPレベルに対してより密接に関係していることを示す。タバコにおけるTSWV-BL Np遺伝子については、少なくとも、植物染色体におけるNP遺伝子の取込部位はウイスル抵抗性で重要でないかも知れないようである。
【0086】
以下の単離株:ブラジル(距離のある関係のウイルス)、T91(密接に関係するウイルス)およびBL(同種単離株)に対する、本発明によるTSWV−BLのヌクレオキャプシド蛋白質遺伝子を含有するトランスジェニックR1およびR2トマトの反応性を決定する実験も行った。これらの実験において、トランスジェニックトマト(エル・エスクレンタム(L.esculentum))は、公表された手法[プラント・セル・リポーツ(Plant Cell Reports)第5巻:81頁(1986年)参照]を用い、トマト黄化壊疽ウイルスのレタス単離株のヌクレオキャプシド蛋白質(N)遺伝子を発芽した子葉にエイ・チュメファシエンス(A.tumefaciens)媒介遺伝子移入することによって産生した。トマト系「Genova80」は形質転換につき選択した。何故ならば、それはTMVに抵抗性を付与し、かくして多数のウイルス−抵抗性系を産生する可能性を生じるTn-22遺伝子を含有するからである。
【0087】
形質転換体をカナマイシン培地で選択し、根が生えたトランスジェニックトマトをポットに入れ、温室に移した。R1およびR2トマト実生はNPT II遺伝子を発現し、これは、植物ゲノムにおけるこの遺伝子の多重挿入を示す。対照的に、18%の実生のみが、検出可能なレベルのN蛋白質を生じた。
【0088】
9つのR1系および3つのR2系を、以下の3つのトスポウイルス、特に、TSWV−BL、TSWV−T91、およびTSWV−Bに対する抵抗性についてテストした。感染性はテスト植物の目による観察に基づくものであった。少しの錆色環または昆虫の損害を除いて植物が健康に見える場合、これらの植物からの抽出物を、ウイルスの存在につきテストするために、エヌ・ベンサミアーナ(N.benthamiana)に対して接種した。以下の表に示すごとく、ほとんどすべての対照トマト植物は、TSWV−BL、TSWV−T91またはTSWV−Bでの接種の3ないし4週間後における植物萎縮、葉の黄色のモザイクおよび皺よりなる典型的な病徴を呈した。しかしながら、R1およびR2トランスジェニック植物の4%のみが、TSWV−BLに感染状態となり、7%がTSWV−T91に感染状態となり、45%がTSWV−Bに感染状態となった。
【0089】
トマト黄化壊疽ウイルス・レタス株の核蛋白質遺伝子を発現する
トランスジェニック・R1またはR2トマトにおけるウイルス抵抗性
接種単離株a
植物系 TSWV-BL TSWV-T91 TSWV-B
R1植物:
T13-1 0/22 1/26 7/24
T13-2 6/20 NTb NT
T13-3 2/42 0/20 12/18
T13-4 0/25 NT NT
T13-9 0/20 NT NT
T13-10 1/50 2/26 11/26
T13-11 0/22 NT NT
T13-12 1/29 NT NT
T13-13 0/22 NT NT
合計 10/252 3/72 30/68
R2植物:
T13-1-7 0/8 2/8 5/8
T13-1-9 0/8 1/8 2/8
T13-1-11 0/8 1/9 5/9
合計 0/24 4/25 12/25
対照 92/95 51/53 52/53
a エヌ・ベンサミアーナ(N.benthamiana)、H423タバコまたはトマトの5-、10-または20-倍希釈葉抽出液で、第一葉ないし第二葉段階の植物を接種した;同一の植物を7日後に再接種し、さらに14日後に病徴を記録した;病徴を呈する植物数/試験植物数として反応性を表した。
b 試験せず
【0090】
従って、前記の記載は、TSWV-BLのN遺伝子を発現するトランスジェニックトマト植物が、TSWV-BL、TSWV-BLに近縁の他のTSWV単離株、ならびにさらに遠縁のTSWV-Bへの感染に対して抵抗性を示すという知見を支持している。
【0091】
さらなる単離株でのさらに限定された実験において、全てのトランスジェニック植物が10W(体菜)単離株に対して抵抗性であったが、対照は感染した。これらの結果は、トランスジェニックトマトが遠縁の単離株よりも近縁の単離株に対してより良好に保護されるということを示している。TSWV-BLのN遺伝子を発現するトランスジェニック・タバコおよびエヌ・ベンサミアーナ(N.benthamiana)とは異なり、N蛋白質の発現レベルは、トランスジェニックトマトにおいて観察された保護と相関がなかった;55%のトランスジェニックトマトが遠縁TSWV-B単離株にも抵抗性であり、このことはトランスジェニック・タバコおよびエヌ・ベンサミアーナ(N.benthamiana)植物においては認められなかった。これらの矛盾は、トマトは本来トスポウイルスに対して感受性が低いということを反映しているのかも知れない。
【0092】
さらに、接種後5および7週の少数の植物においてウイルス分布を評価する実験も行った。各植物の全開葉の小葉から先端半分を摩砕し、エヌ・ベンサミアーナ(N.benthamiana)上に戻し接種した。接種7日後に得た結果は、TSWV-BL.-T91または-B.のいずれかで接種した無病徴トランスジェニック植物のいずれの葉組織からもウイルスが回収できなかったということを示しており、前記に報告した観察知見を追認している。病徴を示すトランスジェニック植物において、ウイルスは植物全体を通して分布していない。例えば、視覚的に終局的に見積もることができないトランスジェニック植物は、8つの葉のうち2つにだけウイルスを含んでいた;植物の地表からの第二葉および先端葉。反対に、感染した対照植物の全ての葉にウイルスが存在し、健康な対照植物においてはウイルスは存在しなかった。
【0093】
ウイルスを脈管系に導入した場合に、抵抗性のトランスジェニック植物が感染させることができるか否かを試験するために、移植接種(graft inoculation)を試みた。1:5、1:10または1:20の希釈率のTSWV-BL、-T91もしくは-Bで接種したR1およびR2植物を、同一の単離株および希釈率にて、感染させた対照植物に移植した。非-トランスジェニック対照が感染したにも拘わらず、34個体のトランスジェニック植物は31日後に無病徴であった。23日後に、トランスジェニック植物の先端46cmを切り取り、新たな成長およびさらなるストレスを誘導した。若く、勢いよく成長している新しい苗芽は接種31日後にいずれの病徴も示さなかったが、接種45日後に各々33%、31%および45%のTSWV-BL、-T91および-Bが葉病徴または茎病徴を示していた。これらの結果は、あるトランスジェニック植物は耐性で、またあるものは感染に対して免であることを示している。
【0094】
かくして、本発明の1つの態様によれば、TSWV-BL単離株のNP遺伝子を発現するトランスジェニック植物は、同種のTSWV-BL単離株および同一血清群の異種単離株(アルカンサスおよび10W体菜)の感染に対して高い抵抗性を示した。さらに重要なことには、抵抗性は他の血清群からベゴニア単離株に至るまで有効である。略言すれば、TSWV-BLの核蛋白質遺伝子を発現するトランスジェニックタバコ植物はTSWVおよびINSVの双方に抵抗性を示し、その保護は遠縁のINSVに対する核蛋白質によって、ならびに同種および近縁のTSWV単離株に対する核蛋白質遺伝子リボヌクレオチド配列によって媒介されているようであることを前記のことは明らかに記載している。異なったTSWV単離株に対して広範囲スペクトル抵抗性の構築植物が示されたのはこれが初めてである。
【0095】
一般的に、外皮蛋白質保護は、免疫ではなくて、感染および病徴発現における遅延および/低下を示すが、本発明は、無病徴でかつ感染ウイルスフリーである有意に高いパーセンテージのトランスジェニック植物を提供する。温室条件下におけるこれらの植物の抵抗性はそれらの生涯を通して保持され、より重要なことは、前記したごとくそれらの子孫にも受け継がれた。
【0096】
たとえあるとしてもほとんどTSWV-BL NPを産生しないトランスジェニック植物が、同種の単離株、およびTSWVの同一血清群内の他の近縁の単離株による感染に高い抵抗性を示すことが本発明において認められた一方で、高レベルのNP遺伝子を発現しているものにおいては保護が認められなかった。
【0097】
TSWVの生物学的多様性は詳細に報告されており、トマトのごとき栽培植物においては遺伝的抵抗性を克服することが報告されている。かくして、多種のTSWV株に対して抵抗性を示すトランスジェニック植物を開発することが極めて重要である。本発明は、この抵抗性を付与するウイルスNP遺伝子を利用してそれが行われるであろうこと、ならびに多様なTSWV単離株にこの抵抗性が示されるであろうことを示している。かくして、TSWV NP遺伝子の発現が種々のTSWV単離株に対して高レベルの抵抗性を付与できるという本発明の知見は、膨大な量の商業的重要性を有する。
【0098】
他の研究のシリーズにおいて、プラスミドBIN19−N+を構築し、実施例IVに従ってアグロバクテリウム・チュメファシエンス(A.tumefaciens)LB4404株に移入し、実施例Vに従いニコチアナ・ベンサミアーナ(Nicotiana benthamiana)に移入した。INSV−Begおよび−LIのヌクレオキャプシド遺伝子をオリゴマープライマーINSV−A(5'−TACTTATCTAGAACCATGGACAAAGCAAAGATTACCAAGG)およびINSV−B(5'−TACAGTGGATCCATGGTTATTTCAAATAATTTATAAAAGCAC)で増幅し、INSV単離体のヌクレオキャプシド遺伝子の5'−コーディング領域および3'−非コーディング領域のそれぞれにハイブリダイズさせた。増幅したヌクレオキャプシド遺伝子断片を実施例IIIに従って精製し、実施例IVに従って消化して配列決定した。
【0099】
合計24のN+(pBIN19−N+で形質転換された)および18N−(ベクターpBIN19で形質転換された)形質転換エヌ・ベンサミアーナ植物を土壌に移し温室で発育させた。全てのN+系列は4−5の葉段階において遺伝子配列を含んでいることがPCRにより確認された。N蛋白質の蓄積の相対レベルはTSWV−BLN蛋白質の抗体を用い、各々独立したR0トランスジェニッククローン系においてDAS−ELISAによって評価した。24のN+系のうち、2つが0.50−1.00のOD405nmの値を有し、17が0.02−0.10の間、残りの5つが0.02より少なかった。健康なエヌ・ベンサミアーナまたはトランスジェニックN−植物は0.02−0/02のOD405nmの読みを与える。R0植物の全てが自家受粉され、以下のトランスジェニック系:(1)N−−2および−6、pBIN19ベクターのみを含む対照トランスジェニック系、(2)N+−28、N蛋白質の検出不可能な量を産生したトランスジェニック系(OD405nm=0.005);(3)N+−21、低レベルのN蛋白質を産生するトランスジェニック系(OD405nm=0.085);および(4)高レベルのN蛋白質を蓄積する2つのトランスジェニック系であるN+−34および−37(OD405nm=0.50−1.00)からの種子を植菌試験のためのカナマイシン選択培地(300μg/ml)上で発芽させた。次いで、これら6つの系をノーザンハイブリダイゼーションによって分析し;N遺伝子転写体の強度はELISA反応のレベルとよく相関していた。
【0100】
6つのR0系からのトランスジェニック実生はカナマイシン選択培地上で種子を発芽させることによって選択し、これらの実生を5種のトスポウイルスで接種した。接種されたR1植物は、もしウイルス病徴が、接種葉に観察されれば感受性と見なした。漏れの可能性を排除するために、トランスジェニック対照N−植物をトランスジェニックN+植物の各々の接種で常に用いた。加えて、各接種抽出物を、まずN+植物を接種し、次いで対照N−植物を接種するのに常に用いた。この一連の研究の結果を以下に示す:
【0101】
エヌ・ベンサミアーナ黄化壊疽ウイルス(TSWV)のヌクレオキャプシド
(N)蛋白質を発現するR1植物の、トスポウイルスでの接種に対する反応性
感染植物数/接種植物数b
TSWV単離株 INSV単離株
R0系 ELISAa BL 10W Beg LI TSWV-B
N--2/-6 <0.02 32/32 32/32 32/32 20/20 32/32
N+-28 0.005 16/16 16/16 15/16 16/16
N+-21 0.085 9/40 17/40 39/40 18/20 40/40
N+-34 0.715 25/28c 28/28 23/28c 28/28
N+-37 0.510 26/28c 22/22 21/28c 16/20c 22/22
a R1植物が由来するR0系のaELISAデータ;
b 感染したエヌ・ベンサミアーナ植物の30倍希釈葉抽出物を3−5葉段階の3枚の植物葉に適用した。各抽出物を常に用いてN+植物を接種した。データは少なくとも接種後2カ月毎日収集し、全身感染植物数/接種植物数として表した;cほとんど全ての感受性R1植物は病徴の出現の顕著な遅延を示すことを示す。
【0102】
上記の表で示したように、対照系N−−2および−6からの全てのR1植物は、試験した全てのウイルスの接種後5−8で全身的な病徴を示した。N+−28系からのR1植物のいずれも検出可能な量のN蛋白質を産生せず、INSV−Begを接種した1の植物を除き全てがこれらのウイルスに感受性であった。接種前に採取したこのN+−28R1植物のリーフディスクのELISA分析により、INSV−Beg抵抗性表現型を保有すると同定された植物が高レベルのN蛋白質を蓄積することが明白に示された(全ての他のN+−28R1植物に対しOD405nmが<0.02であるのと比較しOD405nm=0.78)。
【0103】
N遺伝子を低いレベルで発現しているN+−21系は同種(78%)および近縁のTSWV−10W(57%)単離株に対して良好な抵抗性を示し、は2個のINSV単離株に対しほとんど耐性を示さず(3%および10%);3のN+−21植物のみが、INSV単離株を接種した時に抵抗性の表現型を示した。これらのINSV−耐性N+21 R1植物からの葉の試料は非常に高いELISA反応を示し(OD405nm 0.5ないし1.00)、かくして感受性N+−21植物よりも多量のN蛋白質を与えた(OD405nm 0.02ないし0.50)。N遺伝子の発現が高い系はINSV単離株に対し最も高い抵抗性を示し(18%-5%)、次いで同種TSWV−BL単離株(7%および11%)であり、一方、植物のいずれもTSWL−10Wに対し抵抗性を示さず;しかしながら、INSVまたはTSWV−BLで感染されたN+−34および−37R1植物は病徴の発現において様々な長さの遅れを示す。これらの4つのトランスジェニックN+系からのR1植物のいずれもTSWV−Bに抵抗性でなかった;N+−34および−37系からのR1植物のいくつかは病徴の出現に少しの遅延を示した。
【0104】
N+R1植物におけるN蛋白質産生レベルが異なるトスポウイルスに対する抵抗性と関連しているか否かを決定する実験において、上記表中の接種N+R1植物は、元のR0植物とは関係なく接種前に採取された組織のそのELISA反応の強度に基づき4つの群に再び組織した。N蛋白質を低レベルで発現するN+R1植物(0.02−0.2OD)は、TSWV−BLと−10Wに対し高い抵抗性(100%および80%)を示すが、INSV−Begおよび−LIに対しては全て感受性であり、これは対照のN−植物に対する病徴出現において検出可能な遅延はないことを示す。対照的に、高いレベルのN蛋白質を有するほとんどすべてのN+R1植物(0.20−1.00OD)は、TSWV−BL、INSV−BegおよびLIに対して様々なレベルの保護を示し、病徴の出現の短い遅延から、これらの植物のほとんどが対照N−植物に対する病徴の発展における遅延の様々な長さを示す、完全な耐性の範囲にある。TSWL−10Wに対する高い発現因子において保護は観察されなかった。加えて、N遺伝子発現のレベルに拘らず、N+植物はTSWV−Bに対して抵抗性でなかった;しかしながら、高レベルでN蛋白質を産生するN+R1植物では、短い遅延の病徴の出現が観察された。全ての対照N−R1植物および検出不可能なELISA反応を起こす(0ないし0.02OD)トランスジェニックN+R1植物は試験したすべてのトスポウイルスに対し感受性であった。
【0105】
TSWV−BLヌクレオキャプシド遺伝子を発現するエヌ・ベンサミアーナプロトプラストにおいて遠縁INSVの複製阻害も実験した。これらの実験において、全てのINSV−LIウイルス粒子を用いて3つのトランスジェニック系から単離したプロトプラストを感染させて、形質転換の産物が、いかに入って来るウイルスに影響するかを調べた。ウイルスの複製は、INSV N蛋白質に特異的な抗体を用いてトランスジェニックプロトプラストにおいてINSVを感染するN蛋白質のレベルを測定することによって決定した。DAS−ELISA分析は、与えられた系からの全ての子孫が比較的均一であって、ほとんど全てのR1の子孫が、その親であるトランスジェニック系と類似したトランスジェニックN遺伝子の発現レベルを与えることを示した。これらの結果は、その親系のそれに基づくR1の集団の発現レベルの予測を可能にした。低い発現因子の系N+−21のR1植物由来のプロトプラストはINSV−LIの複製を支持するが、一方、高い発現因子系N+−37のR1植物からのプロトプラストは、低レベルのウイルスの複製が観察される接種の42時間後までこれを支持しなかった。様々な時間間隔(例えば0、19、30および42時間)で同じプロトプラストを、形質転換遺伝子の発現レベルを検査するために、TSWL−BL N蛋白質に特異的な抗体を用いたDAS−ELISA法によっても分析した。予測されるように、N+-21 R1植物からのプロトプラストは比較的低レベル(0.338-0.395,OD405nm)を産生するのに対し、N+−37 R1植物からのプロトプラストは高いレベルを蓄積した(0.822−0.865,OD405nm)。発現レベルは全ての時点で一定であることが判明した。
【0106】
本発明の本態様において、低量のTSWV−BL N蛋白質を蓄積するトランスジェニック・エヌ・ベンサミアーナ植物は同種および遠縁の(TSWV-10W)単離株に高度に対して抵抗性であるが、一方この蛋白質を多く蓄積する植物は同種および遠縁の(INSV−BegおよびINSV−LI)ウイルスの両方に対する適度に保護を有することが示された。より重要なことには、これらの知見は、トランスジェニック・エヌ・ベンサミアーナ植物(INSVの系統的な宿主)はINSV−BegおよびINSV−LI単離株に対し保護されることを示す。
【0107】
上記で示したように、本発明者らは、TSWVのN遺伝子を発現するトランスジェニック植物は、同種単離株に対して抵抗性であり、TSWV−BL遺伝子を発現するかかる植物はTSWVおよびINSVの双方に抵抗性であることを示した。同種および近縁の単離株に対する最良の抵抗性はN蛋白質を低レベルで蓄積しているトランスジェニック植物で見い出されるが、一方TSWV−BL N蛋白質の高レベルを有するトランスジェニック植物は血清学的に遠いINSV単離株により抵抗性であることも示された。この観察によって、本発明者らは、同種および近縁の単離株からの観察される保護における翻訳されたN蛋白質産物の役割に気付き、また植物ゲノムに挿入されたN遺伝子自体か、それともその転写物が保護に関与するかを憶測した。この仮説を試験するために、プロモーターを含まないN遺伝子を含むか、あるいはセンスまたはアンチセンスの翻訳不可能なNコーディング領域を発現するトランスジェニック植物を作出した。発見されたことは、センスおよびアンチセンスの翻訳不可能なN遺伝子RNAは同種および近縁の単離株に対する保護を供給すること、およびこれらのRNA媒介保護は、それぞれのRNA種を低いレベルで合成する植物において最も有効であって、ウイルス複製の阻害を通じて達成されるようである、ということである。
【0108】
より具体的には、トランスジェニック植物に導入されたコーディング配列を図7に示す。示されるように、pBIN19−N構築物は植物形質転換ベクターpBIN19に挿入された無プロモーターN遺伝子を含む(実施例IV参照)。全ての他の構築物は二重のCaMVの35Sプロモーター、アルファルファモザイクウイルスの5'−末端の翻訳リーダー配列およびノパリンシンターゼの3'−非翻訳/ポリアデニル化配列を含む。pBI525は植物発現ベクターであって、この研究においては対照として用いられ;pBI525−mNはN遺伝子の変異(翻訳不能)形を含み;pBI525−asNは翻訳不能N遺伝子のアンチセンス型を含む。変異体N遺伝子の5’末端における1個のヌクレオチドの欠失はダッシュの記号によって示される。ATGコドンに下線を施し、インフレームの終止コドンは太字で示す。
【0109】
実施例VIII
TSWL−BL N遺伝子のプライマー特異的突然変異誘発およびクローニングは以下のごとくに行った:
N遺伝子の全長は、ここに引用して本明細書の一部とみなすファイトパソロジー(Phytopathology)第82巻:1223頁(1992年)に記載されるごとく、逆転写およびポリメラーゼ鎖反応により得た。翻訳不能N−コーディング配列は、オリゴマープライマーA、(AGCATTGGATCCATGGTTAACACACTAAGCAAGCAC)(TSWV−BL N遺伝子の3’−非暗号領域におけるS RNAと同一)、およびB、(AGCTAATCTAGAACCATGGATGACTCACTAAGGAAAGCATTGTTGC)(N遺伝子の5'末端においてS RNAに相補的)を用いたRT−PCRにより同様に作られた。後者のオリゴマープライマーは転写開始コドンの直後のフレームシフト変異および起こり得る翻訳のリードスルーを阻止するためのいくつかの終止コドンを含んでいる。無傷のおよび変異のN遺伝子断片は実施例IIに記載したごとく1.2%アガロースゲルによって精製した。ゲル精製した無傷のおよび変異のN遺伝子断片を適当な制限酵素で消化し、それぞれ実施例IVに記載されているごとく、直接BamHI/およびXbaI消化の植物形質転換ベクターpBIN19およびNcoI消化の植物発現ベクターpBI525にクローンした。得られたプラスミドを同定し、カリフラワー・モザイク・ウイルス35Sプロモーターに対し、無傷の無プロモーターN遺伝子、およびセンスおよびアンチセンスの向きで変異コーディング配列を有するpB1525−mNおよびpB1525−asNを含むpBIN19−Nと命名された。ニコチアナ・タバカム(Nicotiana tabacum)プロトプラストにおける一過性発現分析により発現カセット中の変異Nコーディング配列の翻訳可能性をチェックし;センスまたはアンチセンスの変異Nコーディング配列を含む発現カセットをHindIII/EcoRIによる部分消化(なぜならNコーディング配列は内部のHindIIIおよびExoRI配列を含むので)により切り出し、同酵素で既に切断されている植物形質転換ベクターpBIN19に連結した。pBIN19-Nと同様得られたベクターを、実施例IVに記載した方法を用いてエイ・チュメファシエンスLBA4404株に移入した。エヌ・タバカム・バール・ハバナ・シイブイ(N.tabacum var Havana cv 423)のリーフディスクを、様々な構築物を含有するエイ・チュメファシエンスのLBA4404株で接種し、得られたトランスジェニック植物を自家授粉させ、種子を選択的カナマイシン培地上で発芽させた。
【0110】
PCRは上記のごとく各R0トランスジェニック系で行った。オリゴマープライマーAおよびBを用いてTSWL−BLのN−コーディング配列の存在を決定した。オリゴマープライマー35S−プロモーター(CCCACTATCCTTCGCAAGACCC)をオリゴマープライマーAもしくはBのいずれかと組み合わせて、植物ゲノムに挿入された変異体のNコーディング配列の(CaMV 35Sプロモーターに対する)向きを確認した。DAS−ELISA法を用い、TSWL−BLN蛋白質に対するポリクローナル抗体を用いてトランスジェニック植物においてN蛋白質を検出した。ノーザンブロットによるトランスジェニック植物におけるRNA転写レベルを見積もるために、全植物RNAをナポリ(Napoli)の方法に従って単離し[ザ・プラント・セル(the Plant Cell)第2巻:279頁(1990年)参照]、ホルムアルデヒド含有アガロースゲルで分離した(10μg/レーン)。次いで、該アガロースゲルをエチジウムブロミドで染色して各レーンの全植物RNAの均一性を確認した。ハイブリダイゼーションの条件は製造者によるジーン・スクリーン・プラス(GeneScreen Plus)のプロトコールに記載されている通りである。得られたシグナルブロットを各ブロットに含まれる対照レーンのN遺伝子転写物バンド(高レベルのN遺伝子を産生するmNR1植物)に基づき比較し標準化した。100および150の間の密度値(ヒューレット・スカンジェット・アンド・イメージ・アナリシス・プログラム(Hewlet ScanJet and Image Analysis Program))を示したトランスジェニック植物は高い発現因子と見積もり、一方15および50の間の密度値を有するものは低い発現因子と見積もった。
【0111】
トスポウイルスでのトランスジェニック植物の接種は、特記する以外は前記したごとく行い、接種は3−4葉段階において行った。
タバコ・プロトプラストはR1植物由来の表面を殺菌した葉から調製した
[ツェット・プフランツェフィジオル(Z.Pflanzanphysiol)第78巻:453頁(1992年)に変更を加えたので、参照のこと]単離されたプロトプラスト(6×106プロトプラスト)をPEG法[プラント・アンド・モレキュラー・バイオロジー(Plant Mol.Biol.)第8巻:363頁(1987年)参照]を用いて、OD260nm値0.68の精製したTSWL−BLウイルス粒子抽出物で形質転換した。次いで、形質転換したプロトプラストを暗所にて26℃で終密度1×106プロトプラスト/ml培養培地になるまで培養した。様々な間隔のインキュベーションの後で、培養したプロトプラストをW5溶液で2度洗浄しエンザイム結合緩衝液中、浸透圧のショックにより溶解させた。ウイルスの増殖(複製)はDAS−ELISA法を用いてウイルスのN蛋白質を測定することによって見積もった。
【0112】
記載したごとく、本発明の一の態様は、全く検出できないか僅かしか検出できない量のN蛋白質を生産するトランスジェニック・タバコは同種または近縁の単離株に対し抵抗性であることを示す。この結果は、観察された抵抗性が、入ってきたウイルスN遺伝子と、トランスジェニック植物で産生されたN遺伝子転写物またはNコーディング配列それ自体のいずれかのトランス相互作用によるものであることを示唆する。核N遺伝子の存在が役割を果たすかどうかを試験するために、2つのP0N系からのトランスジェニックP0N R0系およびR1植物を4種のトスポウイルス(TSWV−BL、TSWV−10W、INSV−BegおよびTSWV−B)で攻撃した。無病徴性の植物は抵抗性と評価されたのに対し、何らかの病候を示す植物は感受性であると評価した。全ての接種したR0およびR1植物はウイルスに感受性であった。
【0113】
さらにNトランス遺伝子(transgene)の転写物が保護に関与する可能性を試験するために、センスまたはアンチセンスN遺伝子転写物いずれかを産生するが、N蛋白質は産生しない多数のR0トランスジェニック植物に同種単離株で接種した。結果を以下の表に示す:
【0114】
【表2】
【0115】
接種後7ないし9日で典型的な全身的病徴を示す対照とは異なり、21のmN植物のうち16、および8のasNのうち5のがそのライフサイクルの間中ずっと無病徴であった。接種前に採取された葉の組織のノーザンブロット分析は、全ての抵抗性R0系が低レベルのセンスまたはアンチセンスのN遺伝子RNAを産生するが、一方感受性R0系はRNA種を産生しないかまたは高レベルで産生する。このデータは、トランスジェニック植物のTSWV−BLに対する抵抗性はN遺伝子転写物のその相対的なレベルに関連していることを示唆したので、高いまたは低いN遺伝子転写レベルの4種のmNおよび3種のasN R0系からのトランスジェニックの子孫をカナマイシン含有培地上で発芽させることによって選択した。これらのトランスジェニック植物は、2種のasN系からのいくつかのR1植物は6ないし7葉段階で接種した以外は、3ないし4葉段階において4種のトスポウイルスに対する抵抗性を試験した。結果を以下の表にまとめる:
【0116】
【表3】
【0117】
高い発現因子系mN−2およびmN−7からの全てのR1植物は試験した全てのトスポウイルスによる感染に対して感受性であり、これらの植物は対照と比較して病徴の遅延を示さなかった。対照的に、低い発現因子系mN−13および−18からのR1植物の高い割合のものが同種(TSWV−BL)および近縁(TSWV−10W)単離株に対して抵抗性であったが、遠縁トスポウイルス(INSV−BegおよびTSWV−B)による感染に対しては抵抗性がない。低い発現因子のR0系からのasN R1植物の抵抗性は接種に用いたTSWV単離株によって顕著に影響された。低い発現因子系asN-1および-9からの小さいR1植物(3-4葉段階)の1つを除いて全てが感染したが、同種TSWV−BLまたは近縁TSWV−10Wの単離株を接種した場合は病徴の出現の遅延があった。対照的に、系asN−9からの大きいR1植物(6−7葉段階)のほとんどが両方の単離株に対して抵抗性であった。比較すると、対照R1植物およびasN−4のごとき高発現因子系からのR1植物は、植物の大きさによらずいずれの単離株に対しても抵抗性を示さなかった。アンチセンスRNA媒介される保護は遠縁のINSV−BegおよびTSWV−B単離株による感染に対して有効ではなかった。
【0118】
上記の2つの表に示されたデータの分析によって、センスおよびアンチセンスのRNA−媒介保護はN遺伝子の低い発現因子においてのみ観察されることが示された。高レベルのアンチセンスN遺伝子転写物を産生するR1 asN植物は対照の植物と同程度に感受性があった。対照的に、asNの低い発現因子は、3−4葉段階で接種した場合は病徴の出現に遅延を示し、6−7葉段階で接種した場合はレベルの増加を示した。
【0119】
翻訳不可能Nコーディング配列のセンスまたはアンチセンス型を発現しているタバコ・プロトプラストのウイルス複製の阻害も認められた。この場合には、TSWV−BLの全ウイルス粒子調製物を、トランスジェニック系から単離したプロトプラストをトランスフェクトするのに用いて、入って来るウイルスの複製に与えるセンスまたはアンチセンスのN遺伝子転写物の影響を調べた。ウイルス複製を、トランスフェクトしたプロトプラストに入って来るウイルスのN蛋白質のレベルを測定することによって決定し、高レベルの各RNA転写物を産する植物(mN−7およびasN−4)からのプロトプラストはウイルスの複製を支持し、一方mNの低い発現因子(mN−18)からのプロトプラストは支持しないことが判明した。asNの低い発現因子(asN−9)からのプロトプラストははるかに低いレベルのウイルス複製を支持した。
【0120】
従って、本発明の本態様において、本発明者らは、翻訳不可能N遺伝子コーディング配列のセンスまたはアンチセンス型を発現するトランスジェニック植物は、同種(TSWV−BL)または近縁(TSWV−10W)トスポウイルスには抵抗性であるが、遠縁の(INSV−BegまたはTSWV−B)トスポウイルスには抵抗性でないことを示した。以下の表は、様々な形態のTSWV-BL N遺伝子を発現するトランスジェニック・タバコ間のトスポウイルスに対する抵抗性の比較を示す:
【0121】
【表4】
【0122】
これらの結果により、TSWVでのRNA媒介保護のための本発明の前に述べた態様が確認され、拡大される。さらに、該保護は高レベルのN遺伝子転写物を生産する植物においてよりも、低レベルのN遺伝子転写物を生産する植物において観察され、本明細書中で報告する初期の研究は、高レベルのTSWV−BL N蛋白質を産生するタバコ植物がINSV−Begに対する抵抗を呈することを示すにも拘わらず、この付加的なデータは、INSV−Begに対する抵抗性がセンスおよびアンチセンス型の翻訳不能N遺伝子を発現するトランスジェニック植物では観察されないことを示すので、かくしてINSV−BEGに対する保護はN蛋白質の存在によるものであって、N遺伝子転写物によるものではないことを明白に示唆することを示す。従って、2の異なる機構が、本発明によるTSWVおよびINSVトスポウイルスに対する保護に関与しているように思われる。一の機構はN遺伝子転写物を含み(RNA−媒介)、もう一個はN蛋白質を含む(蛋白質媒介)。加えて、プロトプラスト実験の結果は、N遺伝子のRNA−媒介保護は、ウイルス複製を阻害する過程を通じて達成されることと、上記の表に含まれるデータは、遠縁のINSV−Beg単離株に対する保護はTSWV−BIのN蛋白質によって授与されるのであって、遺伝子転写物によって授与されるのではないことを示唆する。
【0123】
最後に、本発明のもう一個の態様−すなわちトスポウイルス核蛋白質遺伝子の一部はトスポウイルスによる感染に対するトランスジェニック植物の保護を提供するためにさらに研究を行った。N遺伝子RNAは同種および近縁TSWV単離株に対して保護し、一方で、N蛋白質は同種単離株および遠縁のINSV単離株に対し保護すること;N遺伝子のRNE媒介保護は、N遺伝子を低いレベルで発現している植物で有効であるが、一方N蛋白質媒介保護はN蛋白質の高いレベルの蓄積を要すること;およびN遺伝子のRNA媒介保護は、ウイルス複製の阻害を通じて達成されることは上記ですでに示した。この以前のデータに基づき、本発明者らは次に、N遺伝子の一部がウイルスによる感染に対して働くかどうかを決定することに着手した。本発明らは、下記に議論するように、N遺伝子配列の約半分を発現しているトランスジェニック植物はウイルスに対して抵抗性であることを見い出した。
【0124】
以下に、本発明のこの最後の態様を示すためにTSWV−BLの半分のN遺伝子断片のクローニングを記載する。翻訳可能なまたは翻訳不可能なN遺伝子の両方の、第一および第二の半分を、上記に記載したように逆転写、および次いでのPCRによって作成した。図8に示すように、翻訳不可能なN遺伝子断片の半分の5’末端でのヌクレオチドの欠失または挿入はダッシュの記号によって示し;ATGコドンは下線を施し、翻訳不可能なN遺伝子の半分の断片の開始コドンのすぐ後にある可能な終止コドンは太字で示す。
【0125】
N遺伝子の第一の半分はオリゴプライマーi(5'−TACAGTGGATCCATGGTTAAGGTAATCCATAGGCTTGAC)(これはTSWV−BLN遺伝子の中央領域に相補的)および翻訳可能な第一の半分のN遺伝子断片用のii(5'−AGCTAACCATGGTTAAGCTCACTAAGGAAAGCATTGTTGC)、もしくはiii翻訳不可能な第一の半分のN遺伝子断片のための(5'−AGCTAATCTAGAACCATGGATGACTCACTAAGGAAAGCATTGTTGC)を用いてRT−PCRによって生産され、後の方の2つのオリゴマープライマーはN遺伝子の5'末端と同一である。同様に、N遺伝子の第二の半分は、オリゴマープライマーiv(5'−AGCATTGGATCCATGGTTAACACACTAAGCAAGCAC)(これはTSWV−BLN遺伝子の3'非コード領域に相補的)および翻訳可能な半分のN遺伝子断片のためのv(5'−TACAGTTCTAGAACCATGGATGATGCAAAGTCTGTGAGG)、または翻訳不可能な半分のN遺伝子断片のためのvi(5'−AGATTCTCTAGACCATGGTGACTTGATGAGCAAAGTCTGTGAGGCTTGC)を用いてRT−PCRによって生産し、後の方の2個のオリゴマープライマーはN遺伝子の中央領域と同一である。オリゴマープライマーiiiは翻訳コドンのすぐ後にフレームシフト変異および、可能な翻訳のリードスルーを阻害するためのいくつかのインフレームの終止コドンを含む。
【0126】
半分の遺伝子断片を上記のごとく1.2%アガロースゲルで精製し、ゲルで単離した断片を制限酵素NcoIで消化し、NcoI消化した植物発現ベクターpBI525にクローンした。得られたプラスミドを同定し、(1)第一の半分の翻訳可能なN遺伝子を含むpBI525−1N、(2)第一の半分の翻訳不可能なN遺伝子を含むpBI525−1N'、(3)アンチセンスの向きで第一の半分の翻訳可能なN遺伝子を含むpBI525−1N−、(4)第二の半分の翻訳可能なN遺伝子を含むpBI525−2N、(5)第二の半分の翻訳不可能なN遺伝子を含むpBI525−2n'、(6)アンチセンス向きでの第二の半分の翻訳可能なN遺伝子を含むpBI525−2N−と命名した。次いで、発現カセットをHindIII/EcoRIで消化することによってプラスミドpBI525から切り出し、上記のごとく、同じ酵素ですでに切断した植物形質転換ベクターpBIN19に連結した。プラスミドpBIN19と同様得られたベクターを、ホルスターズ(Holsters)(前掲)によって記載された方法を用いてエイ・チュメファシエンス(A.tumefaciens)に移入した。エヌ・ベンサミアーナ(N.benthamiana)のリーフディスクに、様々な構築物を含むエイ・チュメファシエンスのLB4404株を接種した。トランスジェニック植物を自家授粉させ、種子を上記のごとくカナマイシン上で発芽させた。
【0127】
PCRおよびノーザンハイブリダイゼーションPCRによるトランスジェニック植物の分析を、以前記載されたごとく各R0トランスジェニック系において行った。オリゴマープライマーiないしviを、TXWV−BLのNコーディング配列の存在を決定するために用いた。オリゴマープライマー35S−プロモーター(実施例VIII照)を上記のオリゴマープライマーの一つと組み合わせて、植物ゲノムに挿入された半分の遺伝子配列の向き(CaMV35Sプロモーターに対する)を確かめた。ノーザン分析は、実施例VIIIに記載されるごとくに行った。
【0128】
TSWV(TSWV−BL)のレタス単離株を用いてトランスジェニック植物を攻撃した。上記のごとく3−4葉段階のテスト植物を用いて接種した。逃避の可能性を防ぐために、対照の植物を各実験において用い、各接種抽出物を用いてが、まず第一にトランスジェニック植物、次いで対照植物に接種した。
【0129】
本発明の本態様におけるさまざまな構築物を図8に示す。翻訳可能および翻訳不可能な半分のN遺伝子断片をRT−PCRにより合成し、次いで植物発現ベクターpBI525に直接クローンした。RT−PCRによる翻訳不可能な半分のN遺伝子断片の産生のために用いたオリゴマープライマーiiiおよびviは、翻訳開始コドンのすぐ後に変異を有し、得られたリーディングフレームは可能な翻訳のリードスルーを阻害するためのいくつかの終止コドンを含む。従って、第一および第二の半分の翻訳不可能なN遺伝子断片は共に、植物に導入された際に、端を切り取ったN蛋白質断片を産生するのは不可能であるに違いない。次いで、翻訳可能なおよび翻訳不可能な半分のN遺伝子断片を共にセンスまたはアンチセンスの向きでベクターpBI525のCaMV35Sプロモーターの下流に入れた。TSWV−BLの半分のNコーディング配列の発現は、従って、発現ベクターpBI525のアルファルファモザイクウイルス(ALMV)の翻訳されないリーダー配列に融合した二重のCaMV 35Sプロモーターによって制御された。積み重ねられた二重のCaMV 35Sプロモーターを用いる発現ベクターは、一個の35Sプロモーター配列を有する類似のベクターよりも高レベルのmRNA転写を起こすことが公知である。発現カセットをベクターpBI525からパント(pant)形質転換ベクターpBIN19に移入した。対照のプラスミドpBIN19と同様に、得られたプラスミドを次いでエイ・チュメファシエンスLB4404株に移入した。トランスジェニック植物を図8に示すトランスジェニック系の命名と共に得た。
【0130】
全てのカナマイシン抵抗性トランスジェニック系はPCRによって、期待される向きで適切なNコーディング配列を含むことを確認した。種子用に成育させた各トランスジェニックR0系をノーザンブロットにより分析した。6つの1Nのうち6、6つの1N'のうち4、6つの1N−のうち6、6つの2Nのうち6、8つの2N'のうち7、および 7つの2N−トランスジェニックR0のうち6は、半分のN遺伝子RNAを産生することが判明した。
【0131】
トランスジェニックR0植物の一組を同種単離株TSWV−BLで攻撃した。無病徴の植物を抵抗性である評価し、一方でなんらかの病徴(局部の障害または全身的な感染)を示した植物を感受性であると評価した。全ての接種されたR0対照植物はウイルスに感受性であった;対照的に、9つの1N'のうち2、6つの1N−のうち2、10つの2N’のうち4、および8つの2N−R0系のうち一つがウイルス感染に完全に抵抗性であった。1Nおよび2N R0系のどちらとも高レベルの抵抗性を示したが、これらの植物のいくらかは病徴の出現に顕著な遅延を示した。
【0132】
トランスジェニックR0系のもう一組を種子の産生のために成熟させた。実生はカナマイシン含有培地で発芽させ、TSWV−BLを接種した。以下の表に示すように、対照の実生およびいくつかのトランスジェニック系からの実生は単離株に感受性であるのに対し、1N-151、1N'-123、および2N'-134系からの実生は、病徴の発現の遅延から完全な抵抗性まで変化するさまざまなレベルの保護を示した。
【0133】
感染させた植物の数/接種した植物の数
R0系 6DPI 15DPI 30DPI
対照 50/50
1N −149 17/17
1N −151 2/20 13/20 17/20
1N'−123 16/20 17/20 17/20
1N'−124 20/20
1N'−126 19/19
1N−−130 12/15 15/15
1N−−132 18/19 19/19
2N −155 20/20
2N'−134 0/20 10/20 10/20
2N'−135 19/19
2N−−142 20/20
2N−−143 20/20
上記の表において、感染したエヌ・ベンサミアーナ(N.benthamiana)からの
30倍希釈の抽出物を、3−4葉段階のトランスジェニック植物、次いで対照トランスジェニック植物に接種するのに用いた。DPI=接種後の日数
【0134】
要約として、本発明の本態様は、翻訳可能なまたは翻訳不可能なN遺伝子断片の第一のまたは第二のいずれかの半分を発現しているトランスジェニック植物は、同種TSWV−BL単離株に高度に抵抗性であることを示す。この結果は、N遺伝子の一部だけでウイルスに対する抵抗性に十分であることを示す。
【0135】
本発明の上記記載のすべての核酸およびアミノ酸配列のリストは以下の通りである:
【0136】
【0137】
従って、本発明者らは本発明の好ましい具体例を例示し記載したが、本発明の変化および修飾が可能であることが理解されるべきであり、本発明者らはそれゆえ発表された正確な用語を限定することを望むのではなく本発明に様々な用途および状態を付与するためになされる変化および交替を我々に役立たせることを望むものである。このような変化および修飾は、例えば、構造的に類似した核酸配列の置換を含むであろうが、ここにおいて、示される配列および変化した配列間の相違は、もしその変化した配列、即ち、上記記載のものと実質的に類似の結果をうむ配列にほとんど利点をもたらさない。従って、具体的に上記したそれらの配列の機能を実質的に変更することはない塩基(塩基配列中の)またはアミノ酸(ペプチド配列中の)の置換、欠失、挿入あるいは付加は本発明の態様の中に含まれると思われる。加えて、本発明が、本発明に従い植物へのトスポウイルスの感染に対する耐性または免疫を供給する様々な単離株のN遺伝子を一個のカセットにつなぐように修飾されること、およびトスポウイルスに対する広い抵抗性、特にTSWV−BL、TSWV−BおよびINSVに対する抵抗性を供給するためのトランス遺伝子(transgene)としてこのカセットを用いることは本発明者らの努力である。従って、このような変化および交替は本発明と等価である十分な範囲内で適切に意図され、以下の請求の範囲内にある。
その分野、またはそれが最も密接に関連した分野に属するいかなる当業者にも、同じ事をさせ使用させることができるように、十分で、明瞭で、完結で、正確な術語を用いて、本発明およびそこにおいてなされた方法および過程を記載した。
【図面の簡単な説明】
【0138】
【図1】図1は、本発明によるウイルスRNAからNP遺伝子をクローニングするための戦略を示す図である。
【図2】図2は、タバコ・プロトプラストにおける本発明によるトマト黄化壊疽病ウイルスのヌクレオキャプシド蛋白質(NP)遺伝子のイン・ビボ(in vivo)経時的発現を示す図である。
【図3】図3は、本発明によるTSWV-B S RNAにおける、配列決定したcDNAクローンの位置を示す図である。
【図4】図4は、本発明によるTSWV単離株の中での関係を示す樹系図である。
【図5】図5は、本明細書に記載したTSWV単離株の血清学的な関係を示す図である。
【図6】図6は、トランスジェニック植物におけるヌクレオキャプシド蛋白質(NP)の蓄積のレベルとTSWV単離株に対する抵抗性の度合との相関を示す図である。
【図7】図7は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BL N暗号配列を示す図である。
【図8】図8は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BLの半分のN遺伝子断片を示す図である。
【技術分野】
【0001】
トスポウイルス属のウイルスは広範な植物種、特に、タバコ、ラッカセイ、蔬菜および観賞植物に感染する。2種のウイルス種、トマト黄化壊疽ウイルス(TSWV)およびツリフネソウ属壊疽斑点ウイルス(INSV)は、トスポウイルス属に属すると認識されている。
【背景技術】
【0002】
トマト黄化壊疽ウイルス(TSWV)は、核酸-蛋白質複合体がリポ蛋白質エンベロープによって覆われており、かつ唯一のアザミウマ媒介ウイルスである点において植物ウイルスの中では特異である。最近、このウイルスは、ブニヤウイルス科のトスポウイルス属に分類された。TSWVウイルス粒子には、29Kのヌクレオキャプシド蛋白質(「NP」または「N」)、2種の膜関連糖蛋白質(58Kおよび78K)、ならびに恐らくウイルス逆転写酵素のための大きな200K蛋白質が含まれる[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:2207頁(1991年);バイロロジー(Virol.)第56:12頁(1973年);およびジャーナル・ジェネラル・バイロロジー(J.Gen.Virol.)第36巻:267頁(1977年)参照]。そのウイルスゲノムは、L RNA(8900ヌクレオチド)、M RNA(5400ヌクレオチド)およびS RNA(2900ヌクレオチド))と命名される3本の負鎖(-)RNAよりなり[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第36巻:81頁(1977年);ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第53巻:12頁(1981年);およびジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第70巻:3469頁(1989年)参照]、その各々はNPによって包まれている。3種のTSWV単離株からのS RNAの部分配列または全長配列によって、両側センス(ambisense)遺伝子配置を有する2つのオープンリーディングフレーム(ORF)の存在が明らかとなった[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:1頁(1990年)およびジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第72巻:461頁(1991年)参照]。大きな方のオープンリーディングフレームはウイルスRNA鎖上に位置し、52Kの非構造蛋白質をコードする余地を有している。小さな方のORFはウイルスの相補的RNA鎖上に位置し、サブゲノムRNAを通して29KのNPに翻訳される。
【0003】
58Kおよび78Kの膜関連糖蛋白質をコードするオープンリーディングフレームでの両側センス鎖暗号化戦略も、TSWV M RNAに特徴的である。TSWV L RNAは、恐らくはウイルス逆転写酵素のための大きな200K蛋白質をコードすることが配列決定されている。
【0004】
2種のTSWV血清群である「L」および「I」が、構造蛋白質の血清学的分析および細胞病理学的な構造の形態学に基づいて同定され特徴付けられている[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:933頁(1990年)およびファイトパソロジー(Phytopathology)第81巻:525頁(1991年)参照]。それらは、血清学的に保存されたG1およびG2糖蛋白質を有しているが、「I」血清群のNPは「L」血清群のものとは血清学的に異なる。「L」および「I」血清群の間でNPを比較した結果、ヌクレオチドおよびアミノ酸のレベルで各々62%および67%の同一性が示された(ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第72巻:2597頁(1991年)参照]。
【0005】
TSWVは広範な宿主範囲を有し、50科の360植物種を超える植物に感染して、蔬菜および観賞植物に重大な経済的損失を全世界的に引き起こしている。「L」血清群が蔬菜およびタバコのごとき農作物に幅広く認められる一方、「I」血清群は観賞作物にたいてい限定されている。スイカおよび他のウリ科植物に全身感染し、かつそのNPはいずれの血清群のものとも血清学的に関連性がないために、区別される単離株としてウリ科植物単離株が最近同定された[プラント・ディジーズ(Plant Disease)第68巻:1006頁(1984年)参照]。抵抗性植物の育種または非遺伝的方法によってTSWV病の拡大を低下させることが時々できることもあるが、これらの慣用的方法による病害の完全な制御は一般的に困難であることが証明されている[プラント・ディジーズ(Plant Disease)第73巻:375頁(1989年)]。
【発明の開示】
【発明が解決しようとする課題】
【0006】
1986年以来、ウイルスの外皮蛋白質(CP)遺伝子を導入したトランスジェニック植物が該ウイルスによる感染に対してしばしば抵抗性を示すことを、膨大な報告が示してきた。この現象は、通常、外皮蛋白質-媒介保護(CPMP)と呼ばれている。保護の程度は、病徴発現の遅延から病徴およびウイルス蓄積の欠如までに及ぶ。最近の2報の独立した報告[バイオル・テクノロジー(Biol.Tech-nology)第9巻:1363頁(1991年)およびモレキュラー・アンド・プラント-マイクローブ・インタラクションズ(Mol.Plant-Microbe Interact.)第5巻:34頁(1992年)]は、TSWVのヌクレオキャプシド蛋白質(NP)遺伝子を発現するトランスジェニック・タバコ植物が同種の単離株による感染に対して抵抗性を示すことを示した。しかしながら、TSWVは多種の生物的に多様な株に拡散しているため、異なったTSWV単離株による感染に抵抗するトランスジェニック植物の有効性を試験することが非常に重要である。本発明の知見は、本発明によるトランスジェニック植物が、「L」血清群の2種の異種単離株および「I」血清群の1種の単離株に対して抵抗性を示したことを証明することによって、先行の報告のものを超えるものである。本発明者らは、高レベルのNPを蓄積するトランスジェニック植物が「I」血清群の単離株に抵抗性を示す一方、たとえあるとしても非常に低いレベルNPしか蓄積していない植物において、「L」血清群の2種の異種単離株に対する抵抗性が主に認められたことも示す。
【0007】
しかしながら、高レベルのNP蛋白質を蓄積する植物が病徴発現の遅延を示したにも拘わらず、ブラジリアン(Brazilian)単離株に対する抵抗性は認められなかった。TSWV-Bと命名されたこのブラジリアン単離株は、「L」および「I」血清群とは血清学的に区別されるN蛋白質を有し、TSWV-Bがメロンおよびカボチャに全身感染しない点においてウリ科単離株とは生物学的に異なる。
【課題を解決するための手段】
【0008】
従って、本発明の1つの態様は、TSWV-B SのRNAをクローニングし、配列決定して、他のTSWV単離株の公表されている配列と比較することによって該TWV-Bを特徴付けすることである。
【発明を実施するための最良の形態】
【0009】
本発明の種々の態様は、以下の実施例、図面およびデータを包含する本発明の詳細な説明から容易に明らかとなるであろう。
図1は、本発明によるウイルスRNAからNP遺伝子をクローニングするための戦略を示す図である。
図2は、タバコ・プロトプラストにおける本発明によるトマト黄化壊疽病ウイルスのヌクレオキャプシド蛋白質(NP)遺伝子のイン・ビボ(in vivo)経時的発現を示す図である。
図3は、本発明によるTSWV-B S RNAにおける、配列決定したcDNAクローンの位置を示す図である。
図4は、本発明によるTSWV単離株の中での関係を示す樹系図である。
図5は、本明細書に記載したTSWV単離株の血清学的な関係を示す図である。
図6は、トランスジェニック植物におけるヌクレオキャプシド蛋白質(NP)の蓄積のレベルとTSWV単離株に対する抵抗性の度合との相関を示す図である。
図7は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BL N暗号配列を示す図である。
図8は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BLの半分のN遺伝子断片を示す図である。
【0010】
さらに詳細には、図2は、ポリエチレングリコール(PEG)を用いてタバコ葉肉プロトプラストに構築物が移入されたNP遺伝子の経時的発現を図示している。続いて、NP遺伝子を発現させるためにその形質転換プロトプラストを2日間インキュベートした。プロトプラストから蛋白質を抽出し、TSWV NPに対する抗体を用いた二重抗体サンドイッチ酵素結合免疫吸着アッセイ(DAS-ELISA)によってNPについて試験した。NP−およびNP+は、各々プラスミドpBI525-NP−およびpBI525-NP+で形質転換したプロトプラストを表す。コーティング用の抗体濃度は5μg/ml、酵素結合物の希釈率は1:250である。データは、基質添加後30、60および90分に収集した。
【0011】
図3において、5つの重複cDNAクローンは、TSWV-BのS RNA地図以下の尺度で示されている。これらのクローンは、TSWV-Bに感染したエヌ・ベンサミアーナ(N.benthamiana)から単離した二本鎖RNAからのランダムプライマーで合成した。
【0012】
図4において、配列は、GCGシークエンス・アナリシス・ソフトウェア・パッケージ(GCG Sequence analysis software package)のパイルアップ・プログラムを用いて比較した。水平線は遺伝子距離に比例するが、垂直線は任意の長さであり、有意性はない。
【0013】
さらに詳細には、図5において、エヌ・ベンサミアーナ・ドミン(N.benthamianaDomin)をTSWV単離株[TSWV-BL(レタス単離株)、アルカンサス(Arkansas)、10W体菜(pakchoy)(TSWV-10W))、ベゴニアおよびブラジル(TSWV-B)]で感染させた。感染リーフディスク(0.05g)を12mlの酵素結合緩衝液中にて摩砕し、TSWV-BLビリオン(BL ビリオン)、またはTSWV-BLのNP(BL-NP)もしくはTSWV-IのNP(I-NP)に対して生起させた抗体を用いるDAS-ELISAにより分析した。コーティング用の抗体濃度は1μg/ml;BLウイルス粒子については結合物の希釈率は1:2000、BL-NPについては希釈率は1:250、I-NPについては希釈率は1:1000であった。基質添加後10分(BL)、50分(BL-NP)または30分に結果を収集した。
【0014】
図6に関しては、TSWV-BLのNPに対して生起させた抗体でNP蓄積につきDAS-ELISAでトランスジェニック植物をアッセイした。植物を基質添加後150分に判読し、該トランスジェニック植物を4つのカテゴリーにグループ分けした:0.050より低いOD405nm値、0.050ないし0.200の間のOD405nm値、0.200ないし0.400の間のOD405nm値、および0.400よりも高いOD405nm値。対照NP(-)植物のOD405nm判読値は0ないし0.05であった。同一の植物をアルカンサス(Ark)および10W体菜(10W)単離株またはベゴニア単離株で攻撃し、各植物の感受性を接種約12日後に記録した。結果は、アルカンサスおよび10Wチュウゴクキャベツ単離株で接種した51個体のR1 NP(+)植物、およびベゴニア単離株で接種した139個体のR1 NP(+)植物から収集した。バー上の数字は、試験したR1 NP(+)植物の合計数を表す。
【0015】
実施例I
TSWV-BL RNAの単離:
TSWV-BL単離株は、ダチュラ・ストラモニウム・エル(Datura stramonium L.)から以下のごとく精製した:感染組織を3倍容量の緩衝液(0.033MのKH2PO4、0.067MのK2HPO4、0.01MのNa2SO3)と共にワーリング・ブレンダーで45秒間摩砕した。そのホモジネートを前記緩衝液で湿らせた4層のチーズクローズを通して濾過し、7,000rpmにて15分間遠心した。そのペレットを組織の元の重量に等しい量の0.01M Na2SO3に再懸濁し、8,000rpmにて15分間再度遠心した。その後に、上清を元の組織重量の1/10に等しい量の0.01M Na2SO3に再懸濁した。そのウイルス抽出液を9,000rpmにて15分間遠心し、その上清を、0.01MのNa2SO3中に作成した10-40%スクロース段階勾配上に注意深く負荷した。23,000rpmにて35分間遠心した後に、ウイルスゾーン(メニスカスより約3cm下)を採取し、2倍容量の0.01M Na2SO3で希釈した。その半-精製ウイルスを27,000rpmにて55分間ペレット化した。
【0016】
実施例II
TSWVおよびウイルスRNAの精製
TSWV-BL単離株[プラント・ディジーズ(Plant Disease)第74巻:154頁(1990年)]は、実施例Iに記載のごとく、ダチュラ・ストラモニウム・エル(Datura stramonium L)から精製した。0.04%のベントナイト、10μg/mlのプオテイナーゼK、0.1Mの炭酸アンモニウム、0.1%(w/v)のジエチルチオカルバミン酸ナトリウム、1mMのEDTA、および1%(w/v)のドデシル硫酸ナトリウム(SDS)の溶液中に精製したウイルスを再懸濁し、65℃にて5分間インキュベートして、直ちにH2O-飽和フェノールから抽出し、続いてさらにクロロホルム/イソアミルアルコール(24:1)から抽出した。ウイルスRNAを2.5容量のエタノール中に沈殿させ、蒸留水に溶解した。
【0017】
実施例III
cDNAおよびPCRに基づくNP遺伝子のクローニング:
第一鎖cDNAは、グブラー(Gubler)およびホフマン(Hoffman)により記載されているランダムプライマーを用いて、精製したTSWV-BLのRNAから合成した[ジーン(Gene)第25巻:263頁(1983年)参照]。第二鎖は、RNアーゼ H/DNAポリメラーゼで試料を処理することにより作製した。得られた二重鎖cDNA試料を、スクロース濃度勾配遠心法によりサイズ-分画し、EcoRIメチラーゼによりメチル化し、EcoRIリンカーを付加した。EcoRIで消化した後に、子ウシ腸アルカリ性ホスファターゼで処理することによってその5'-末端リン酸基が除去されているpUC18のEcoRI部位に、cDNA試料を連結した。イイ・コリ(E.coli)DH5αコンピテント・セル(ベセスダ・リサーチ研究所(Bethesda Research Laboratories))を形質転換し、TSWVのcDNAインサートを含むクローンを、50μg/mlのアンピシリン、IPTGおよびX-galを含有する寒天培地上に平板培養することによって一次選抜した。アルカリ溶解法を用いて、選抜クローンからプラスミドDNAを単離し[ビイアールエル・フォーカス(BRL Focus)第11巻:7頁(1989年)参照]、EcoRI制限酵素消化に続くジーンスクリーン・プラス(GeneScreen Plus)・ナイロンフィルター(デュポン社(DuPont))上へのDNAトランスファーによってインサートのサイズを測定した。TSWV-BL S RNAのcDNAインサートを含むプラスミドクローンは、以下に記載するごとく、TSWV-CPNH1 S RNAのヌクレオチド配列(GCAAGTTCTGCGAGTTTTGCCTGCT)に相補的な32P-標識オリゴマー(AGCAGGCAAAACTCGCAGAACTTGC)に対するハイブリダイゼーションによって同定した[ジャーナル・オブ・ジェネラル・バイロロジー(J.Gen.Virol.)第71巻:001頁(1990年)参照]。幾つかのクローンをアガロースゲル上で同定し分析して、インサートのサイズを測定した。クローンpTSWVS-23は、約1.7kbの長さの最も大きなcDNAインサートを含んでいることが判明した。
【0018】
ポリメラーゼ鎖反応(PCR)を使用することにより全長NP遺伝子を得た。第一鎖cDNA合成は、TSWV NP遺伝子の5'末端(TSWV-CPNH1のヌクレオチド2571ないし2773位)においてS RNAに相補的である(TSWV-10Wのヌクレオキャプシド遺伝子を合成するのにも用いた)オリゴマープライマーJLS90-46(5'->3')AGCTAACCATGGTTAAGCTCACTAAGGAAAGCを用いて、20μl反応混合液中、37℃にて30分間で行った。その反応混合物には、1.5μlのウイルスRNA、1μgのオリゴマープライマー、0.2mMの各dNTP、1×PCR緩衝液(ジーンアンプ・キット(GeneAmp kit)、パーキン-エルマー-シータス社(Perkin-Elmer-Cetus))、リボヌクレアーゼ・インヒビター中の20UのRNA(プロメガ社(Promega))、2.5mMのMgCl2、および25UのAMV逆転写酵素(プロメガ・コーポレイション(Promega Corporation))が含まれていた。95℃にて5分間加熱することによって反応を終了させ、氷上にて冷却した。次いで、各1μgのオリゴマープライマーJLS90-46およびJLS90-47(5'->3')、(TSWV-10Wのヌクレオチド遺伝子を合成するのにも用いた)(後者のオリゴマーは該遺伝子の3'非コーディング領域(TSWV-CPNH1の1919ないし1938のヌクレオチド位)のS RNAと同一である)を用いる業者の取り扱い説明書(パーキン-エルマー-シータス社(Parkin-Elmer-Cetus))に従って、10μlのcDNA/RNAハイブリッドを用いてNP遺伝子をPCR増幅させた。典型的なPCRサイクルは、92℃にて1分間(変性)、50℃にて1分間(アニーリング)、および72℃にて2分間(重合)であった。試料は、1.2%アガロースゲル上に直接負荷して分離させた。分離したNP遺伝子断片をアガロースゲルから抽出し、エタノール沈殿し、20μlの蒸留水に溶解した。
【0019】
実施例IV
植物発現ベクターおよび植物形質転換ベクターの構築
実施例IIIからゲル単離したNP遺伝子断片を、50μl反応緩衝液[50mMのトリス-HCl(pH8.0)、10mMのMgCl2、0.1MのNaCl]中の制限酵素NcoIで37℃にて3時間消化し、NcoIで消化した植物発現ベクターpB1525に直接クローン化した。得られたプラスミドを同定し、カリフラワー・モザイク・ウイルス(CaMV)35Sプロモーターに対してセンス向きであるプラスミドをpB1525-NP+と命名し、逆向きであるものをpB1525-NP−と命名した。パン(Pang)らにより記載されているごとく[ジーン(Gene)、第112巻:229頁(1992年)参照]、ニコチアナ・タバカム(Nicotiana tobacum)プロトプラストにおけるNP遺伝子の経時的発現により、この発現カセットのNP産生能を測定した。次いで、(NP遺伝子は内部HindIIIおよびEcoRI部位を含んでいるため)HindIII/EcoRIでの部分消化によって、NP遺伝子を含有する発現カセットをpB1525-NP+から切り出し、同酵素で切断した植物形質転換ベクターpBIN19(クロンテック・ラボラトリーズ,インコーポレイティド(Clontech Laboratories,Inc.))に連結した。ホルスターズ(Holsters)らにより記載されている方法[モレキュラー・アンド・ジェネラル・ジェネティクス(Mol.Gen.Genet.)第163巻:181頁(1978年)参照]を用いて、得られたベクターpBIN19-NP+および対照プラスミドpBIN19をエイ・チュメファシエンス(A.tumefaciens)LBA4404株に移入した。
【0020】
クローンpTSWV-23およびPb1525-NP+中のインサートのヌクレオチド配列分析は、ジデオキシ法、T7ポリメラーゼ(ユウエス・バイオケミカルズ社(U.S.Biochemicals)、シークエナーゼ(Sequenase)TM)、ならびにシーミニアック(Siemieniak)らにより記載されている二重鎖配列決定法[アナリティカル・バイオケミストリー(Analyt.Biochem.)第192巻:441頁(1991年)参照]を用いて測定した。ヌクレオチド配列は双方のDNA鎖から決定し、この情報をジェネティクス・コンピューター・グループ(ジイシイジイ(GCG),ワイオミング州・マジソン(Madison,WI))から販売されているコンピューター・プログラムを用いて、TSWV単離株CPNH1の公表されている配列と比較した。
【0021】
タバコ・プロトプラストにおけるNP遺伝子の経時的発現も調製した。大規模アルカリ法(large scale alkaline method)を用いて、クローンpTSWVS-23および(PCR-構築NP遺伝子インサートを含んでいる)pUC18cpphasTSWV-NPについてのプラスミドDNAを単離した。NcoI消化によってPCR-構築NP遺伝子インサートをクローンpBIN25-NP+から切り出して、販売されている配列決定用のフランギング・オリゴマー・プライマーを利用した。発現カセットpUC18cpphasは、ファゼオラス・ブルガリス(Phaseolus vulgaris)種子貯蔵遺伝子であるファゼオリン由来のポリ(A)付加シグナルをそれが利用する点を除いて、pUC18cpexpに同様である。これらのプラスミドDNAを、ベックマンTi 70.1固定角ローターを用いて、2つのCsCl-エチジウムブロミド勾配分画に付した。DNA配列は、前記したジデオキシ法および二重鎖プラスミドDNA配列決定法を用いて得た。ヌクレオチド配列反応物を1m長のサーモスタット制御された(55℃)配列決定用ゲル上で電気泳動し、平均約750bpのヌクレオチド配列の読みを得た。ヌクレオチド配列は、双方のクローン化インサートの双方のDNA鎖から決定して、精度を確認した。TSWV-BL S RNA単離株からのヌクレオチド配列情報を、以下に論じるごとく、コンピューター・プログラム(ジイシイジイ(GCG)、ワイオミング州・マジソン(Madison,WI))を用いてTSWV単離株CPNH1およびL3と比較した。
【0022】
TSWV-BL S RNAのクローン化cDNAおよびPCR-構築インサートのヌクレオチド配列および予想アミノ酸配列ならびに、それらとTSWV-CPHN1 S RNAのヌクレオチド配列との比較を以下に示す。シーミニアック(Siemieniak)の二重鎖ジデオキシ配列決定法を用いてTSWV-BL S RNAクローンpTSWVS-23(TSWV-23)およびpBI1525-NP+(TSWV-PCR)のヌクレオチド配列を得、ジーンバンク(GeneBank)登録番号D00645に報告されているTSWV-CPNH1 S RNAのヌクレオチド配列の関連領域と比較した。TSWV-CPNH1 S RNAのヌクレオチド配列は、ド・ハーン(De Haan)(1990年)により報告されており、以下の配列によって表される:
【0023】
【化1】
【0024】
TSWV-CPNH1 S RNA上の非構造蛋白質遺伝子の不完全な予想アミノ酸配列は、1位の核酸で始まり783位で終了する核酸コドンで終了し、これを以下に掲げる:
【0025】
【化2】
【0026】
以下に図示するTSWV-23のヌクレオチド配列は、前記で得られたTWSV配列と酷似しており、非構造遺伝子の1/2とヌクレオキャプシド蛋白質遺伝子の1/2を含んでいる。
【0027】
【化3】
【0028】
以下に図示する本発明によるTSWV-PCRの核酸配列も、前記で得られたTSWV配列と酷似しており、ヌクレオキャプシド蛋白質遺伝子の全体をカバーしている。
【0029】
【化4】
【0030】
合わせて、クローン化TSWV-23インサートはTSWV-PCRインサートと重複しており、合わせて、それらは本発明によるTSWV-BL S RNAの2028個のヌクレオチドを表している。本発明によるこの2028個のヌクレオチド配列は、非構造遺伝子の一部分とヌクレオキャプシド蛋白質遺伝子の全体を含んでいる。合わせた配列は以下の通りである:
【0031】
【化5】
【0032】
この比較により、クローンpTSWVS-23のcDNAインサートが約760bpの52K蛋白質のウイルス成分遺伝子、完全な遺伝子間(intergenic)領域(492bp)、および450bpのNP遺伝子(NP遺伝子の約半分)を含んでいることが示された。このクローン化インサートは、EcoRI認識部位に正確に位置するその3'-末端を有しており、このことはcDNAクローニング工程の間の不完全なEcoRIメチル化を示している。このクローンは完全なTSWV-BLのNP遺伝子を含んでいないにも拘わらず、PCR遺伝子構築NP遺伝子の配列と450bPの重複をそれが有するため(TSWVにつき、全2028bpのTSWV-BL S RNAがヌクレオチド配列中に示されている)、その配列はかなり重要であった。このTSWV-BL PCR-構築およびTSWV-CPNH1 NP 遺伝子の間の配列比較により、合計21ヌクレオチドの相違(2.7%)、およびその内の8個がアミノ酸置換(3.1%)をコードしているが明らかとなった。突然変異が導入されることが知られているTaqポリメラーゼを用いてこのPCR構築NP遺伝子を得たため、これらの相違のうち幾つかはPCR増幅の間に導入された可能性がある。しかしながら、これらのヌクレオチド相違のうち15個は、TSWV-BL cDNAおよびPCRクローンの間の重複領域内に位置し、これらのヌクレオチド相違の1つ(TSWVの1702位;TSWV-PCRの485位)を除く全ては、双方のTSWV-BL S RNA起源のクローンによって共に保有されている。この比較は、これらの2つのクローン化NP遺伝子領域のヌクレオチド配列間の差異に全くそうではないにせよ大いには寄与しないことを明らかに示した。1702位におけるヌクレオチドの差異の結果、IleとSerとのアミノ酸置換が起こり、この差異でさえTSWV-BL単離株内の相同性の欠如によるものであろう。
【0033】
実施例V
アグロバクテリウム媒介形質転換:
アグロバクテリウム株LAB4404(クロン・テク(Clon Tech))の液状培地中に一晩浸漬することによって、ベクターpBlN19−NP+または対照プラスミドpBlN19を含有する該アグロバクテリウム株と共にニコチアーナ・タバカム・バール・ハバナ・シイブイ(Nicotiana tabacum var Havana cv)423の葉ディスクをインキュベートし、該接種したリーフディスクを非選択的MS培地上で3日間インキュベートした[サイエンス(Science)第227巻:1229頁(1985年)参照]。形質転換した細胞を選択し、苗条再生のために、300μg/mlカナマイシンおよび500μg/mlカルベニシリンを含有するMS培地中で再生させた。小植物を無ホルモン培地に移した後、根を誘導した。根が出た形質転換体を土壌に移し、温室条件下で成長させた。該MS培地は、十分な強度のMS塩(シグマ社(Sigma))、30g/lのグルコース、1mg/lのBAおよび1mlのB5ビタミン[1mg/mlのニコチン酸、10mg/mlのチアミン(HCl)、1mg/mlのピリドキシン(HCl)、100mg/mlのMyo−イノシトール]を含有する。トランスジェニック植物を自家受粉させ、種子をカナマイシン培地上で選択的に発芽させた。
【0034】
実施例VI
蛋白質の血清学的検出:
二重抗体サンドイッチ酵素結合免疫吸着法(DAS−ELISA)を用いて、TSWV−BL NPに対するポリクローナル抗体でトランスジェニック植物におけるNP遺伝子の発現を検出した。各試料は、酵素結合緩衝液[リン酸緩衝セーライン、0.05%ツイーン(Tween)20、2%ポリビニルピロリドン40、および0.2%オバルミン]の3ml中で、植物の頂部第2葉からのリーフディスク(約0.05g)を摩砕することによって調製した。タバコ・プロトプラストについては、遠心後の細胞抽出物をアッセイで直接用いた。トランスジェニック植物およびタバコ・プロトプラスト双方からの試料の10倍および3倍希釈物をDAS−ELISAの直前に作成した。
【0035】
ウェスタンブロットについては、2×SDS/試料緩衝液(0.126M トリス緩衝液、20%グリセロール、2%SDS、2%2−メルカプトエタノール、および0.01mg/mlブロモフェノールブルー)の0.25ml中でリーフディスク(約0.05g)を摩砕した。該ホモジネートを遠心し、負荷の前に上清を沸騰させた。蛋白質(10〜20μl試料/レーン)を分離し、膜上にブロットした。次いで、製造業者のimmunoselectキット指示マニュアル(ジブコ・ビイアールエル・ライフ・テクノロジーズ・インコーポレイテッド(Gibco BRL Life Technologies Inc.))に従って該膜を加工した。全ウイルス粒子に対する抗体を健康なタバコ植物からの細胞抽出物で予め吸収させ[プラント・ディジーズ(Plant Disease)第70巻:501頁(1986年)参照]、2μg/mlの濃度にてウェスタンブロットで用いた。
【0036】
TSWV−BLウイルス粒子、またはTSWV−BLもしくはTSWV−1に対して生起させた抗体を用い、TSWV単離株(TSWV−BL、アルカンサス、10W体菜、ベゴニアまたはブラジル)の血清学的反応をDAS−ELISAで検定した。
【0037】
実施例VII
TSWV単離株でのトランスジェニック植物の接種
接種物は、ニコチアナ・ベンタミアナ・ドミン(Nicotiana benthamiana Domin.)を異なるTSWV単離株で感染させ、緩衝液(0.003M KH2PO4、0.067M K2HPO4および0.01M Na2SO3)15ml中でエヌ・ベンタミアナ植物の感染させた葉(0.5g)を摩砕する(接種1ないし2週間後)ことによって調製した。接種抽出物を、トランスジェニック植物のコランダム散布葉上で直ちに擦り、接種した葉を引き続いて水で濯いだ。TSWVは摩砕後イン・ビトロでは高度に不安定であるので、接種物の各バッチを用いて、NP遺伝子を含有するNP(+)植物をまず接種し;各接種物の最後の接種植物は、常に、ベクター配列単独を含有する対照NP(-)植物であって、特定のウイルス接種物はなお接種の最後において感染性であることを確実とした。
【0038】
接種7〜15日後に、局所病巣および全身感染についてのデータを採り、特に示す以外は、接種した植物の数にわたる全身感染した植物の数として以下の表に表した。この表において、「ELISA」の下で収集したデータは、それからR1植物が由来するR0系のデータであり;ベゴニア単離株はR1植物に局所病巣を誘導し、および抵抗性は、接種した植物の数にわたる局所病巣生成植物の数として表し;NTはテストをしなかったことを示す。
【0039】
ウイルス(TSWV)をスポットしたタバコのヌクレオキャプシド蛋白質
(NP)遺伝子を発現するR1植物のTSWV単離株での接種に対する反応
TSWV単離株に対する反応
エライザ
(R0pl.) BL アルカンサス 10W体菜 ベゴニア ブラジル
R0系
NP(+) 0.015 0/10 4/25 3/24 29/40 36/36
NP(+)4 0.386 6/30 21/23 18/21 9/48 42/42
NP(+)9 0.327 0/20 NT 20/20 - -
NP(+)14 0.040 0/20 - 9/20 8/18 18/18
NP(+)21 0.042 0/15 5/15 3/15 2/4 6/6
NP(+)22 0.142 0/20 - 15/20 31/36 36/36
NP(+)23 0.317 0/20 - 16/20 - -
NP(-) - 42/42 24/24 62/62 66/66 54/54
【0040】
前記したごとく、TSWVのS RNA成分中に存在するTSWV−BL NP遺伝子の単離株は、2つの戦略を用いてアプローチした。cDNAクローニング戦略は、TSWV−CPNH1 S RNAに相補的なオリゴマープローブに対するハイブリダイゼーションによって同定したところによると、TSWV−BL S RNAに由来するcDNAインサートを含有するいくつかのクローンを生じた。クローンpTSWVS−23は、約1.7kb長さにマップされる最長インサートを含有していた。第2の戦略は、TSWV−CPNHl S RNAの公表された配列、および直接全TSWV−BL RNAからの発現のためにNP遺伝子を増幅し、作成するPCRを利用した。オリゴマープライマーJLS90−46および−47を合成し、JLS90−46はNP遺伝子の5’暗号領域中のS RNAに相補的であり(TSWV−CPNH1の2051〜2073位)、一方、JLS90−47はNP遺伝子の3’非暗号領域のものである(TSWV-CPNH1の1218〜1237位)。両プライマーは、引き続いてのクローニングのための制限酵素Nco1についての認識部位を含有し、プライマーJLS90−46は植物コンセンサス翻訳開始コドン配列(AAXXATGG)を有し、これは、翻訳に際して、NP遺伝子の第3コドン(GTT)に翻訳開始コドンを融合させることが期待された。翻訳開始コドンのTSWV−BL NP遺伝子の第3コドンへの融合を行って、いずれのアミノ酸コドンも取り込むことなくNcoI認識部位を保持した。かくして、PCTで作成したTSWV NP遺伝子の発現は、天然NPよりもN末端で2のアミノ酸(Ser-Lys)だけ短いTSWV-BL NPを生じるであろう。
【0041】
この特異的に増幅させた約850bpのDNA断片をNcoIで消化し、植物発現ベクターpB152Sにクローンした。CaMV 35Sプロモーターに対するTSWV−BL NP遺伝子の向きは、制限酵素部位マッピング(EcoRI、HindIII、AvaIおよびAiwN1)によって決定した。適当な向きでインサートを含有するいくつかのクローンを単離し(pB1525−NP+)、他のものは反対の向きでインサートを含有した(pB1525−NP−)。また、この制限酵素部位マッピングは、クローンpB1525−NP+のインサートが、TSWV-CPNH1 NP遺伝子で見い出されるものと同一の制限酵素部位を含有することを示した。かくして、TSWV−BL NP遺伝子の発現は、発現ベクターpB1525のアルファルファモザイクウイルス(ALMV)の5’−非翻訳リーダー配列に融合した二重CaMV 35Sプロモーターによって制御された。重ねた二重CaMV 35Dプロモーター要素を利用する発現ベクターは、単一の35Sプロモーター要素を利用する同様のベクターよりも高レベルのmRNA転写を生じる。
【0042】
3つのpB1525−NP+クローンを一時的にタバコ・プロトプラストで発現させて、増幅されたDNA断片がNPをコードすることを確認した。これを達成するために、PEG法によってクローンをタバコ・プロトプラストに移入し、インキュベーションの2日後、全TSWV−BLウイルス粒子に対する抗体を用い、DAS−ELISAによって発現されたNPを検出した。プラスミドpB1525-NP+中にNP遺伝子を保有するタバコ・プロトプラストで高レベルのNPが産生され:一方、アンチセンスNP配列で形質転換したタバコ・プロトプラスト(pB1525−NP−)ではNPは検出されなかった。
【0043】
前記したごとく、クローンpB1525−NP+のPCR構築インサートおよびクローンpTSWV−23のtehcDNAインサートを二重鎖配列決定に付した。cDNAおよびPCRクローンの配列解析により、各々、1.71kbおよび865bpのインサートが明らかにされ、これは、配列TSWV−CPNH1 S RNAと比較した場合、クローンpTSWV−23のcDNAインサートが52K蛋白質ウイルス粒子成分遺伝子、完全な遺伝子間領域(492bp)およびNP遺伝子の450bp(該遺伝子の約半分)を包含することを示す。このクローン化インサートはEcoRI認識部位に正確に位置するその3’末端を有し、これは、cDNAクローニング間における不完全なEcoRIメチル化を示唆する。このクローンは完全なTSWV−BL NP遺伝子を含有しないにも拘わらず、その配列は非常に重要である。というのは、それはPCR構築NP遺伝子の配列と450bpの重複を有するからである。このTSWV−BL PCR構築NP遺伝子とTSWV−CPNH1 NP遺伝子の間の配列の比較は、合計21個のヌクレオチドの差異(2.7%)を明らかにし、そのうち8つはアミノ酸置換をコードする(3.1%)。このPCR−構築NP遺伝子は、突然変異を取り込むことが知られている、Taqポリメラーゼを用いて得られたので、これらの差異のうちいくらかはPCR増幅の間に導入することができる。しかしながら、これらのヌクレオチド差異のうち15はTSWV−BL cDNAクローンおよびPCRクローンの間の重複領域内に位置し、これらの差異のうちの1つ(1702位)を除いたすべてが、両TSWV−BL R RNA由来クローンに存在する。この比較は、明らかに、PCR増幅は、これらの2つのNP遺伝子のヌクレオチド配列間の差異に全てではないにせよ大いには寄与しないことを示す。1702位におけるヌクレオチドの差異の結果、IleのSerでのアミノ酸置換が起こり、この差異でさえTSWV−BL単離株内における相同性の欠如によるであろう。
【0044】
ヌクレオチドの差異がTSWV単離株間の発散に帰されるという可能性は、
TSWV−CPNH1、TSWV−L3、およびTSWV−B1 S RNAの間の他の配列決定された領域との比較によって支持される。これらの比較は以下の表に示す:
【0045】
【表1】
【0046】
CPNH1およびL3単離株からのNP遺伝子のヌクレオチド配列は相互に3.1%だけ相違し、BL単離株とは、ほとんど同程度(2.5%)相違する。しかしながら、CPNH1およびBL単離株の間のNPアミノ酸配列は、L3およびBLの間、またはCPNH1およびL3単離株の間よりも、かなり相違する。また、前記表の結果は、これらのTSWV単離株のNP遺伝子領域が、52K蛋白質のアミノ酸配列間の差異が7.9ないし10.6%の範囲であるので52K蛋白質よりも高い程度の選択圧に付されており、NPのアミノ酸配列について見い出されているよりも2倍以上であることを明らかにする。ヌクレオチド配列の発散は遺伝子間領域の間で最大であり、これは、この領域が遺伝子領域よりも低い選択圧に付されてことを示す。
【0047】
トランスジェニック植物におけるNP遺伝子配列の存在は、まず、PCR分析によって確認された。NP遺伝子を挟む配列に相同なプライマーを用い、約800bpのNP DNA断片がトランスジェニックNP(+)植物の全DNAから特異的に増幅され、一方、対応する断片は対照NP(-)植物では検出されなかった。NP遺伝子の発現はDAS−ELISAによって各R0トランスジェニック植物でアッセイし、結果は以下の表に示す:
【0048】
トマト黄化壊疽ウイルス(TSWV)のヌクレオキャプシド蛋白質(NP)遺伝子を発現するR0トランスジェニック植物のTSWV-BL単離株での接種に対する反応
植物年齢 R0クローン ELISAa 病巣/葉b NP(+):NP(-)c
7〜8葉 NP(+)1 0.374 7(199) 1:28
NP(+)2 0.015 0(199) 0:199
NP(+)3 0.407 23(102) 1:4
NP(+)4 0.386 2(102) 1:51
NP(+)5 0.023 0(124) 0:124
NP(+)6 0.197 35(325) 1:9
NP(+)7 0.124 1(325) 1:325
9〜10葉
NP(+)8 0.344 36(36) 1:1
NP(+)9 0.327 2(20) 1:10
NP(+)10 0.406 34(33) 1:1
NP(+)11 0.156 5(20) 1:4
NP(+)12 0.133 9(57) 1:6
NP(+)13 0.144 2(7) 1:4
NP(+)14 0.040 0(19) 0:19
NP(+)16 0.053 0(10) 0:10
5〜6葉
NP(+)20 0.487 203(117) 2:1
NP(+)21 0.042 0(117) 0:117
NP(+)22 0.142 0(208) 0:208
NP(+)23 0.317 223(208) 1:1
NP(+)24 0.051 0(35) 0:35
NP(+)25 0.286 13(35) 1:3
NP(+)26 0.037 0(22) 0:22
NP(+)27 0.425 305(22) 14:1
a トランスジェニック植物におけるNPの産生は二重抗体サンドイッチ酵素結合免疫吸着法(DAS−ELISA)によって検定した;コーティング用のウイルス粒子に対する抗体の濃度:1μg/ml;TSWV−BLへのコンジュゲートの希釈:1:250;基質添加後150分に採取した結果;405nmにおける読み
b 接種した葉で発達した局所病巣は接種7日後に計数した。データは3つの接種した葉の平均を表す。括弧に入れたデータは、同一接種物で接種した対照NP(+)植物から生じた病巣の数である。
c pBIN19−NP+
で形質転換したNP(+)植物で発達した局所病巣-対-同一接種物で接種した場合の対照NP(-)植物で発達した局所病巣の比率
【0049】
23のNP(+)クローンのうち、10は高レベルのNPを産生し、5は中レベルのNPを蓄積し、および残りの8は低レベルのNPを産生した。トランスジェニック植物で発現されたNPのサイズはウェスタンブロット法によって分析した。タバコ抽出物からの多くのポリペプチドは、抗体を健康なタバコ植物からの抽出物で予め吸収させたにも拘わらず、全ウイルス粒子に対する抗体に対して反応性であった。それらのうち、1つのバンドのみが、NP遺伝子で形質転換したタバコ植物からのポリペプチドのパターンに対してユニークであった。このポリペプチドは約29kDaであると評価され、これは、天然NPの予測されるサイズに近い。同様のサイズの抗体反応性蛋白質バンドは、ベクターpBIN19を含有するトランスジェニック植物からの抽出物では見い出されなかった。
【0050】
TSWV−BL単離株でのタバコ葉の接種の結果、天候条件および植物の生理学的段階に応じて、全身感染または壊死性局所病巣いずれかとなるであろう。R0植物をウイルス抵抗性につきTSWV−BLでテストした場合、TSWV−BLは、接種6〜8日後に、対照NP(-)植物の接種した葉上に典型的な壊死性病巣を誘導した。しかしながら、トランスジェニックNP(+)植物は、対照NP(-)植物に比して、ウイルスに対する抵抗性スペクトルを示した。23のNP(+)植物のうち7つはいずれの局所病巣も発生せず、あるいは発生した病巣の数は対応する接種したNP(-)植物上のものよりも少なくとも20倍小さかった。3つのNP(+)植物は中程度の反応を有していたが(対照よりも5−ないし19−倍小さい)、残りの9の植物は低い抵抗性であるか、または全く抵抗性がなかった。接種したNP(+)またはNP(−)植物のいずれも全身感染を示さなかった。無病徴R0植物をそのライフサイクルの最後までモニターし、そのライフサイクルを通じて病徴が観察されなかった。無病徴NP(+)植物の接種した葉を、シイ・キノラ(C. quinola)植物の葉上のウイルスの存在についてチェックした。高度に抵抗性のNP(+)植物のTSWV−BL−攻撃葉からウイルスは回収されず、これは、該ウイルスがこれらのNP(+)植物では複製もしくは増殖できないことを示す。
【0051】
選択したR0植物からのリーフディスクを継代クローンし、再生した小植物を該ウイルスによって攻撃した。すべての継代クローンしたR0植物は、それらの対応するオリジナルのR0植物と同様のレベルの抵抗性を示した。
【0052】
TSWVは広く分布して、多くの生物学的距離の株が存在するので、異なるTSWV単離株による感染に抵抗性を示すトランスジェニック植物の効果もテストした。5つのTSWV単離株を本実験で選択して、カナマイシン含有培地上で発芽したR1植物を攻撃した:TSWV−BL、アルカンサス、10W体菜、ベゴニアおよびブラジル。最初の3つの単離株は全ウイルス粒子およびTSWV-BL(通常のTSWV「L」血清群)に対する抗体に対して反応性であった(図5参照)。ベゴニア単離株はTSWV−IのNP(「I」血清群)に対する抗体と強く反応したが、TSWV−LP NPに対して生起したものに対しては反応せず、従って、「I」血清群に属した。ブラジル単離株の、TSWV−BLまたはTSWV−I血清群のNPに対する抗体に対する検出可能な反応は見い出されず、それは、TSWV−BLの全ウイルス粒子に対する抗体には弱い反応性であった。さらに、この単離株は感染したタバコおよびエヌ・ベンサミアーナ(N.benthamiana)の葉上で全身の斑および皺を引き起こしたが、カボチャまたはキュウリを感染せず、これは、それがウリ科植物単離株とは区別される単離株であることを示す。これらの結果は、この単離株が第3の血清群であると考えられることを示す。
【0053】
7つのR0系に由来する実生をカナマイシン培地上で発芽させ、前記したTSWV単離株で接種した。感染性データは接種7日後に開始し、毎日記録した。TSWV−BL、アルカンサス、10W体菜またはブラジル単離株で接種した植物は、もしウイルス病徴が未接種葉で観察されれば、感受性とした。ベゴニア単離株で接種した植物は、局所病巣が接種葉で観察されれば、感受性とした。というのは、この単離株はタバコで全身性感染を引き起こさないからである。すべての接種した対照NP(-)R1植物は、これらの5つの単離株による感染に対して感受性であった。それらは、ベゴニアで接種したトランスジェニックR1植物が接種葉で局所病巣を生じたに過ぎないことを除き、接種12日後に全身感染した。しかしながら、ほとんどすべてのNP(+)R1植物は、同種単離株TSWV-BLに対して高い抵抗性を示したが、かなり低いパーセントのNP(+)R1植物が異種単離株アルカンサス、10W体菜およびベゴニアに対して抵抗性であった。他方、系NP(+)4からのNP−高発現NP(+)R1植物のいくつかでは、病徴性発現においてわずかな遅延(1ないし2日)が観察されたにも拘わらず、7つのトランスジェニック系からのすべてのNP(+)R1植物はブラジル単離株に対して感受性であった。
【0054】
抵抗性R1植物は、そのライフサイクルを通じて無病徴のままであった。17のより低い病徴性のNP(+)植物の接種した葉を、チェノポジウム・キノア(Chenopodium quinoa)植物の葉上の逆接種によってウイルスの存在についてチェックした。無病徴NP(+)植物の接種した葉からウイルスは回収されず、これは、該ウイルスがこれらのNP(+)植物で複製もしくは増殖できないであろうことを示す。
【0055】
トランスジェニック植物におけるNPの蓄積レベルと異種TSWV単離株に対する抵抗性の程度との間の関係も調べた。
【0056】
前記したデータの分析は、低レベルNPのR0系に由来するR1植物は「L」血清群(アルカンサスおよび10W体菜)の異種単離株に最良の抵抗性を付与し、一方、高レベルNPのR0系からのR1は「I」血清群に属するベゴニア単離株に抵抗性であることを示す。例えば、低NP発現系NP(+)2、14および21からの接種したR1植物の平均76%はアルカンサスおよび10W体菜による感染に対して抵抗性であり、一方、これらの単離株に対する抵抗性は、高NP発現系NP(+)4、9および23からの同様に接種した植物の11%のみで観察された。他方、ベゴニア単離株は、低NP発現系NP(+)2、14および21からのR1植物の79%に感染したが、高NP発現系NP(+)4からのR1植物では19%のみに感染した。
【0057】
従って、低レベルのNP遺伝子を発現するトランスジェニックR1植物は単離株10W体菜(「L」血清群)での感染に対して高度に抵抗性であるが、ベゴニア単離株(「I」血清群)での感染に対しては抵抗性でなかったと結論された。対照的に、高NP発現R1植物はベゴニア単離株による感染に対して非常に抵抗性であったが、10W体菜からの単離株による感染に対しては抵抗性でなかった。
【0058】
かくして、異種単離株に対して抵抗性と個々の植物におけるNP発現との関係を正確に定量するのは興味深かった。本明細書中で報告する多数の接種実験において、トランスジェニック植物の葉試料はアルカンサスおよび10W体菜単離株での接種前に採取した。また、NP発現レベルと抵抗性との間の明らかな関係の観察を行った後に、ベゴニア単離株で接種した植物の非接種葉から試料を採取した。サンプリングの後者の方法は、ベゴニア単離株による感染からの干渉なくして行うことができた。何故ならば、この単離株はタバコにおいて全身感染を引き起こさないのみならず、TSWV−BL NPに対する抗体とも反応しないからである。単離したTSWV−BLのNPに対して生起した抗体を用い、DAS−ELISAによって、すべての試料を相対的NPレベルにつき検定した。図5および6はトランスジェニックR1植物(それらが由来するR0系を問わない)におけるNPレベルとアルカンサスおよび10W体菜単離株に対する、またはベゴニア単離株に対するそれらの抵抗性との間の関係を示す。非常に低いまたは検出不可能なELISA反応性(0〜0.05 OD405nm)を有するほとんどすべてのトランスジェニックR1植物はアルカンサスおよび10W体菜単離株(「L」血清群)による感染に対して抵抗性であるが、ベゴニア単離株(「l」血清群)に対しては感受性であった。対照的に、高ELISA反応性(0.4〜1.0 OD405nm)を与えるほとんどすべてのR1植物はベゴニア単離株に対して抵抗性であったが、アルカンサスおよび10W体菜単離株に対して感受性であった。
【0059】
ブドウ葉巻ウイルスで感染させた組織からのdsRNAの単離で首尾よく用いられたことがある方法[アクタ・ホルティカルチャー(Acta Horticulture)第186巻:51頁(1986年)、およびカン・プラント・ディス・スルブ(Can. Plant Dis. Surv.)第68巻:93頁(1988年)]の組合せを用い、TSWV−Bで感染させたエヌ・ベンサミアーナ(N.benthamiana)植物から二本鎖(ds)RNAを単離した。該dsRNAをcDNA合成につき選択した。というのは、この単離株からのウイルス粒子の単離は可能ではなかったからである[プラント・ディジーズ(Plant Disease)第74巻:154頁(1990年)参照]。TSWV-BのS RNAに特異的なcDNAライブラリーを作成するために、二本鎖S RNAをゲル精製し、メチル水銀処理によって変性し、ランダムプライマーを用いてプロメガ社によって提供されたcDNA合成手法に付した。合成したcDNA断片をEcoRIアダプターを介してEcoRI消化のλ ZAPII(ストラタジーン社(Stratagene)) にクローン化し、ゲル精製S RNAの逆転写によって調製したcDNAプローブを用いるコロニーハイブリダイゼーションによって陽性クローンを同定した。数ダースの陽性クローンをアガロースゲル上で分析し、ほとんど全部のTSWV−B S RNAをカバーする、最長インサート(L1、L22およびL30)を含有する3つの重複クローンのみを選択した(図3参照)。
【0060】
クローンL1、L22およびL30中のインサートのヌクレオチド配列は、双方のDNA鎖から、最初はユニバーサルおよび逆プライマーによって、次いでTSWV−BのS RNAの配列決定のために設計された内部プライマーによって決定した。配列決定はサンガー(Sanger)のジデオキシ法、T7ポリメラーゼ(ユウエス・バイオケミカルズ(U.S.Biochemicals)、シークエナーゼ(Sequenase)TM)、およびシーミニアック(Siemieniak)によって記載された二本鎖の配列を決定する方法(アナリティカル・バイオケミストリー(Analyt.Biochem.)参照)を用いて行った。これらのクローンの配列解析は、それぞれ、1.994kb、2.368kbおよび1.576kbのインサートを示し、これらの配列はS RNAゲノムの93%を示した(図3参照)。組み立てた配列をジェネティクス・コンピューター・グループ(Genetics Computer Group)(ジイシイジイ、ウィスコンシン州(GCG,Madison,WI))から販売されているコンピューター・プログラムを用いてTSWV単離株CONH1、L3、IおよびBLの配列と比較することにより解析した。
【0061】
コンピューター分析によって、2.842kbの組み立てた配列は完全な52K非構造蛋白質遺伝子、完全な遺伝子間領域(629bp)、および737bpのNP遺伝子(該NP遺伝子の39のN−末端タクレオチドのみが表されなかった)をカバーすることが示された。N遺伝子のこの失われた領域を得るために、N遺伝子の開始コドンからの62個のヌクレオチド配列と同一のプライマーTTCTGGTCTTCTTCAAACTCAをポリヌクレオチドキナーゼで末端標識して前記したcDNAライブラリーをスクリーニングした。5つの推定クローンが得られた。5つのクローンの配列解析は、クローンS6およびS7のみがN遺伝子のこれらの39の失われたヌクレオチドを含有することを示した。また、後者のクローンはS RNAの3’極末端を包含した。
【0062】
S RNAの5’極末端は、5’RACEシステム(ジブコ(GIBCO))を用いて得られた。TSWV−BのssRNAおよびTSWV−Bで感染させたタバコ植物から単離した全RNAを共に用いて、te TSWV−B S RNAのヌクレオチド746−763位に相補的なオリゴヌクレオチド(5'-CTGTAGCCATGAGCAAAG)を持つ第1鎖cDNAを合成した。ターミナルデオキシヌクレオチジルトランスフェラーゼを用いて第1鎖の3'−末端にdCTPをテールした。次いで、ホモポリマーテールにアニールするアンカープライマー、およびTSWV−B S RNAのヌクレオチド512−529位にアニールするオリゴヌクレオチド(5'-TTATATCTTCTTCTTGGA)を用い、PCRによって、テールしたcDNAを増幅した。該PCR増幅断片をゲル精製し、配列解析のために、T−ベクターpT7Blue(ノバゲン(Novagen))に直接クローン化した。8つの独立したクローンを、S RNAの5’領域(TSWV−B S RNAのヌクレオチド40−57位)に密接なオリゴマープライマー(5'-GTTCTGAGATTTGCTAGT)で配列決定した。得られたクローンのうち6つはS RNAの5’極末端を含有し、これらのクローンからの5'-末端ヌクレオチド配列は同一であった。かくして、TSWV−B S RNAの完全なヌクレオチド配列は3049ヌクレオチド長であった。
【0063】
かくして、3つの従前に配列決定したクローンと共にこれらの2つのクローン(L1、L22、L30、S6およびS7)は、前記した全3032ヌクレオチドをカバーしていた。TSWV−CPNH1およびTSWV−1の末端配列との比較により、18ヌクレオチドの5'極末端は組み立てられた配列中には表されていないにも拘わらず、TSWV−B S RNAの3’−極末端はTSWV-I S RNAの3'-極末端と同一であり、TSWV−CPNH1の3'−極末端とは15ヌクレオチド異なることが明らかとなった。TSWV単離株間の末端配列の保存は、ブニヤウイルス属の他のメンバーの保存し一致し、これは、末端配列は安定な塩基対合構造を形成し、それはその複製およびキャプシド化に関与し得るという仮説を支持する。
【0064】
本発明によるTSWV−B(前記したブラジリアン単離株)のS RNAゲノムの完全なヌクレオチド配列は以下の通りである。
【0065】
【化6】
【0066】
【化7】
【0067】
本発明による非構造(前記単一下線)およびヌクレオキャプシド蛋白質の帰結されるアミノ酸配列は以下の通りである。
【0068】
【化8】
【0069】
および
【0070】
【化9】
【0071】
前記したヌクレオキャプシド蛋白質遺伝子はウイルスの相補鎖上にあり、TSWV−Bのヌクレオキャプシド蛋白質遺伝子は以下の通りである。
【0072】
【化10】
【0073】
TSWV−Bの完全なS RNAは3049ヌクレオチド長であるべきであり、TSWV−CPNH1のS RNAよりも134ヌクレオチド長い。この差異はTSWV−B S RNAの長い遺伝子間領域に主として帰せられる。TSWV−B S RNAの配列決定した領域の解析により、前記した2つのオープンリーディングフレームが明らかとされ、これは他のTSWV単離株と同様である。より大きい方は、ヌクレオチド88に始まり、ヌクレオチド1491で終わるウイルスRNA鎖上に位置していた。ウイルス相補鎖上のより小さい方は、ヌクレオチド2898における開始コドンおよびヌクレオチド2122の終止コドンによって境界が決せられた。2つのオープンリーディングフレームは629ヌクレオチドの遺伝子間領域によって分離されていた。TSWV−B S RNAの配置したヌクレオチドおよびアミノ酸配列と他の単離株のそれとのパーセント相同性比較を示すところの、全配列決定したTSWV−B S RNAと以下の表中の他の単離株のS RNA領域との比較は以下の通りである。
【0074】
全53K蛋白質遺伝子 遺伝子間 29K蛋白質遺伝子
比較a nt nt aa nt nt aa
B/CPNH1 76.4b 80.0 86.1(78.3)c 72.4 77.5 91.5(79.1)
B/L3 75.8 79.0 89.0(82.0) 76.4 78.0 91.1(79.9)
B/BL 76.3 - - 72.8 77.6 90.3(79.5)
B/I 63.0 - - - 63.1 69.7(55.3)
CPNH1/L3 94.8 95.6 92.0(89.4) 89.2 96.8 99.6(98.5)
CPNH1/BL 96.4 - - 95.9 97.2 98.8(96.9)
CPNH1/I 62.7 - - - 60.8 69.5(55.1)
L3/BL 95.1 - - 92.6 97.3 99.2(98.5)
L3/I 60.9 - - - 60.9 69.5(55.1)
I/BL 61.7 - - - 60.9 68.8(53.9)
a 単離株TSWV−CPNH1(2.916kb)、TSWV−L3(2.837kb)、TSWV−BL(2.037kb)およびTSWV−1(1.144kb)の部分的および完全なS RNA配列を、TSWV−BのS RNA配列(3.049kb)との比較のために用いた。
b パーセント類似性は、GCG配列決定分析ソフトウェアパッケージのプログラムBESTFITを用い、それらのヌクレオチドまたは予想されるアミノ酸配列の比較によって計算した。
c パーセント同一性は括弧に入れた。
【0075】
前記したごとく、最大のヌクレオチド配列の類似性(75.8%−76.4%)はL−タイプの単離株(CHNH1、L3およびBL)で示された。より程度が低いが、TSWV−B S RNA とI血清群に帰属させられたTSWV−IのS RNAの間にヌクレオチド配列類似性(63%)があった。比較すると、L-タイプの単離株(CHPN1、L3およびBL)の配列決定したS RNA領域は94.8%−96.4%のヌクレオチド配列類似性を保有していた。
【0076】
777ヌクレオチドのオープンリーディングフームは予測される分子量が28700Daの258アミノ酸のN蛋白質をコードする。TSWV単離株からのNオープンリーディングフレームの配列比較により、TSWV単離株CPNH1、L3およびBLからのN遺伝子のヌクレオチド配列は、それらが相互に異なる(2.7%−3.2%)よりも、TSWV−Bとはかなり(22%−22.5%)異なっていることが明らかにされた。免疫学的分析の結果と一致して、CPNH1、L3およびBL単離株のNアミノ酸配列は、TSWV−Bに対する(90.3%−91.5%の類似性または79.1%−79.9%の同一性)よりも、相互に対してより密接である(98.8%−99.6%の類似性または96.9%−98.5%の同一性)。かなり低い相同性が、ヌクレオチド(63.1%)およびアミノ酸(69.7%の類似性または55.3%の同一性)レベル双方においてTSWV−Iに対して観察された。262アミノ酸をコードするTSWV−IのNオープンリーディングフレームを除き、他の単離株のNオープンリーディングフームは258個のアミノ酸についてコードする。コンピューター解析は、TSWV−I Nオープンリーディングフームの余分な残基はアミノ酸配列の挿入の結果であることを示す(残基82ないし84および残基116)。1つの可能なN−グリコシル化部位が残基68に見い出される。
【0077】
1404ヌクレオチドの第2のオープンリーディングフレームは、分子量が52566Daと予測される467個のアミノ酸の非構造蛋白質をコードする。TSWV−CPNH1およびTSWV−L3の相同オープンリーディングフレームの比較は、ヌクレオチドレベルで80%および79%の類似性、ならびにアミノ酸レベルで86.1%(または78.3%の同一性)および89%(または82.0%の同一性)の類似性を示した。このオープンリーディングフレームは4つの可能なグリコシル化部位を含有し、これはTSWV-CPNH1およびTSWV-L3と正確に同一の位置に存在する。
【0078】
いくつかの挿入のため、TSWV−B S RNAの遺伝子間領域は、TSWV-CPNH1およびTSWV−L3の対応物よりも各々126および41ヌクレオチド長かった。プログラムFOLDによる配列解析は、遺伝子間領域は、当該遺伝子間領域のA−リッチのストレッチと共にU−リッチのストレッチを内部的に塩基対合させることによって、非常に複雑で安定なヘアピン構造を形成することができ、これは、最小自由エネルギー値によって表してTSWV−CPNH1およびTSWV−L3から得られたものと同様の安定性を有することを示した。この内部塩基対合構造は転写終始シグナルとして働き得る。
【0079】
また、前記表の結果は、TSWV−BのN蛋白質は52K蛋白質よりも高い選択圧に付されていることが明らかとなった;52K蛋白質のアミノ酸配列の間の類似性はNPのアミノ酸配列で見い出されるものよりも低い。ヌクレオチド配列の発散は遺伝子間領域のうちで最も高く、これは、この領域がいずれの遺伝子領域よりも高い選択圧に付されていることを示す。
【0080】
TSWV−Bおよび他の4つのTSWV単離株の間の発展的な関係を解析し、これを図4に示すが、そこでは、発展的な系統樹組織はこれらのTSWV単離株について収集された血清学的データの関連性と合致する。かくして、本発明におけるTSWV−Bは、I−タイプの単離株TSWV−Iに対するよりもL−タイプの単離株に対してより密接であるが、L−タイプの単離株が相互に対するよりもL−タイプの単離株に対して類似性がかなり低い。
【0081】
病徴性発現のわずかな遅延にも拘わらず、トランスジェニック植物はTSWVのブラジル単離株に対して抵抗性を示さなかった;血清学的結果は、この単離株が「L」および「I」タイプの単離株から区別され、ウリ科植物単離株から生物学的に異なることを示す。かくして、ブラジル単離株は、なお、TSWVのもう1つの血清群に属するであろう。いずれにせよ、感染性の結果は、単一のNP遺伝子がトスポウイルス属におけるすべての単離株に対する抵抗性を提供するようである。
【0082】
低いまたは検出できないELISA反応性(0−0.05 OD405nm)を与える本発明によるトランスジェニック植物は、「L」血清群の異種単離株(アルカンサスおよび10W体菜)による感染に対して抵抗性であり、一方、これらの単離株に対する保護は高レベルのNPを蓄積する植物では見い出されなかった。対照NP(−)植物のELISAの判読値(0.05 OD405nm)に比較して、これらのトランスジェニック植物はもしあってもほとんどTSWV−BL NPを生産しないであろう。同様の結果がトランスジェニック植物で観察されており、そこでは、CPの蓄積は検出されず:これらはウイルス感染に対して高い抵抗性を示した。この現象を基礎づける機構は現在知られていない。このタイプの抵抗性は、恐らくは、攻撃ウイルスのマイナスセンス複製RNAにハイブリダイズさせ、必須の宿主因子(例えば、レプリカーゼ)に結合するかあるいはウイルス粒子組立体に干渉することによって、トランスジェニック植物で生じるCP RNA分子とウイルス複製との干渉に帰すことができるようである。
【0083】
しかしながら、同種TSWV−BL単離株に対する抵抗性は、明らかに、NP遺伝子の発現レベルから独立していることに注意されたい。TSWV−BLで接種した個々のR1植物の相対的NPレベルは測定しなかったが、これらの接種R1植物(合計145のテスト植物)で生じたNPは検出不可能ないし高度の範囲であると仮定するのは合理的である。
【0084】
「L」血清群の異種単離株からの保護の場合とは対照的に、TSWV−I血清群のベゴニア単離株からの保護が高NP発現R1植物で見い出された。「L」血清群のNPヌクレオチド配列と「I」血清群のそれとの比較により、ヌクレオチドおよびアミノ酸における各々62%および67%の同一性が明らかにされた。2つの血清群のNP遺伝子の差異は余りにも大きくて、トランスジェニック植物で生じたNP(「L」血清群)は、「I」血清群のベゴニア単離株を攻撃するに際して機能不全蛋白質として働くであろう。この「欠陥」外皮蛋白質のウイルス粒子への取込みは、ウイルスの運動またはそのさらなる複製を阻害する欠陥ウイルスを生じるであろう。このタイプの相互作用は、保護には高レベルのNPを要すると予測される。また、ベゴニア単離株に対する抵抗性は、R1植物で産生されるNP転写体の、ウイルス複製での干渉を含む。もしこれが真実であれば、異種ウイルスの複製を阻害するのに、(2つのNP遺伝子の異種性質のため)より多くのNP転写体を要するであろう。
【0085】
「L」および「I」血清群の異種単離株に対する抵抗性に対する個々のR1植物におけるNPレベルの関係を示す結果について明らかな説明はないが、これらは明らかな傾向であると考えられる。というのは、データは多数(190)の植物から得られたものだからである。かくして、個々の植物におけるCPまたはNPレベルの測定はNPまたはCPレベルを抵抗性に関係付けるためのより正確な方法を提供すると考えられる。この形式のデータ解析によって、抵抗性は、それらが由来するR0系のNPレベルに対するよりも、各テスト植物におけるNPレベルに対してより密接に関係していることを示す。タバコにおけるTSWV-BL Np遺伝子については、少なくとも、植物染色体におけるNP遺伝子の取込部位はウイスル抵抗性で重要でないかも知れないようである。
【0086】
以下の単離株:ブラジル(距離のある関係のウイルス)、T91(密接に関係するウイルス)およびBL(同種単離株)に対する、本発明によるTSWV−BLのヌクレオキャプシド蛋白質遺伝子を含有するトランスジェニックR1およびR2トマトの反応性を決定する実験も行った。これらの実験において、トランスジェニックトマト(エル・エスクレンタム(L.esculentum))は、公表された手法[プラント・セル・リポーツ(Plant Cell Reports)第5巻:81頁(1986年)参照]を用い、トマト黄化壊疽ウイルスのレタス単離株のヌクレオキャプシド蛋白質(N)遺伝子を発芽した子葉にエイ・チュメファシエンス(A.tumefaciens)媒介遺伝子移入することによって産生した。トマト系「Genova80」は形質転換につき選択した。何故ならば、それはTMVに抵抗性を付与し、かくして多数のウイルス−抵抗性系を産生する可能性を生じるTn-22遺伝子を含有するからである。
【0087】
形質転換体をカナマイシン培地で選択し、根が生えたトランスジェニックトマトをポットに入れ、温室に移した。R1およびR2トマト実生はNPT II遺伝子を発現し、これは、植物ゲノムにおけるこの遺伝子の多重挿入を示す。対照的に、18%の実生のみが、検出可能なレベルのN蛋白質を生じた。
【0088】
9つのR1系および3つのR2系を、以下の3つのトスポウイルス、特に、TSWV−BL、TSWV−T91、およびTSWV−Bに対する抵抗性についてテストした。感染性はテスト植物の目による観察に基づくものであった。少しの錆色環または昆虫の損害を除いて植物が健康に見える場合、これらの植物からの抽出物を、ウイルスの存在につきテストするために、エヌ・ベンサミアーナ(N.benthamiana)に対して接種した。以下の表に示すごとく、ほとんどすべての対照トマト植物は、TSWV−BL、TSWV−T91またはTSWV−Bでの接種の3ないし4週間後における植物萎縮、葉の黄色のモザイクおよび皺よりなる典型的な病徴を呈した。しかしながら、R1およびR2トランスジェニック植物の4%のみが、TSWV−BLに感染状態となり、7%がTSWV−T91に感染状態となり、45%がTSWV−Bに感染状態となった。
【0089】
トマト黄化壊疽ウイルス・レタス株の核蛋白質遺伝子を発現する
トランスジェニック・R1またはR2トマトにおけるウイルス抵抗性
接種単離株a
植物系 TSWV-BL TSWV-T91 TSWV-B
R1植物:
T13-1 0/22 1/26 7/24
T13-2 6/20 NTb NT
T13-3 2/42 0/20 12/18
T13-4 0/25 NT NT
T13-9 0/20 NT NT
T13-10 1/50 2/26 11/26
T13-11 0/22 NT NT
T13-12 1/29 NT NT
T13-13 0/22 NT NT
合計 10/252 3/72 30/68
R2植物:
T13-1-7 0/8 2/8 5/8
T13-1-9 0/8 1/8 2/8
T13-1-11 0/8 1/9 5/9
合計 0/24 4/25 12/25
対照 92/95 51/53 52/53
a エヌ・ベンサミアーナ(N.benthamiana)、H423タバコまたはトマトの5-、10-または20-倍希釈葉抽出液で、第一葉ないし第二葉段階の植物を接種した;同一の植物を7日後に再接種し、さらに14日後に病徴を記録した;病徴を呈する植物数/試験植物数として反応性を表した。
b 試験せず
【0090】
従って、前記の記載は、TSWV-BLのN遺伝子を発現するトランスジェニックトマト植物が、TSWV-BL、TSWV-BLに近縁の他のTSWV単離株、ならびにさらに遠縁のTSWV-Bへの感染に対して抵抗性を示すという知見を支持している。
【0091】
さらなる単離株でのさらに限定された実験において、全てのトランスジェニック植物が10W(体菜)単離株に対して抵抗性であったが、対照は感染した。これらの結果は、トランスジェニックトマトが遠縁の単離株よりも近縁の単離株に対してより良好に保護されるということを示している。TSWV-BLのN遺伝子を発現するトランスジェニック・タバコおよびエヌ・ベンサミアーナ(N.benthamiana)とは異なり、N蛋白質の発現レベルは、トランスジェニックトマトにおいて観察された保護と相関がなかった;55%のトランスジェニックトマトが遠縁TSWV-B単離株にも抵抗性であり、このことはトランスジェニック・タバコおよびエヌ・ベンサミアーナ(N.benthamiana)植物においては認められなかった。これらの矛盾は、トマトは本来トスポウイルスに対して感受性が低いということを反映しているのかも知れない。
【0092】
さらに、接種後5および7週の少数の植物においてウイルス分布を評価する実験も行った。各植物の全開葉の小葉から先端半分を摩砕し、エヌ・ベンサミアーナ(N.benthamiana)上に戻し接種した。接種7日後に得た結果は、TSWV-BL.-T91または-B.のいずれかで接種した無病徴トランスジェニック植物のいずれの葉組織からもウイルスが回収できなかったということを示しており、前記に報告した観察知見を追認している。病徴を示すトランスジェニック植物において、ウイルスは植物全体を通して分布していない。例えば、視覚的に終局的に見積もることができないトランスジェニック植物は、8つの葉のうち2つにだけウイルスを含んでいた;植物の地表からの第二葉および先端葉。反対に、感染した対照植物の全ての葉にウイルスが存在し、健康な対照植物においてはウイルスは存在しなかった。
【0093】
ウイルスを脈管系に導入した場合に、抵抗性のトランスジェニック植物が感染させることができるか否かを試験するために、移植接種(graft inoculation)を試みた。1:5、1:10または1:20の希釈率のTSWV-BL、-T91もしくは-Bで接種したR1およびR2植物を、同一の単離株および希釈率にて、感染させた対照植物に移植した。非-トランスジェニック対照が感染したにも拘わらず、34個体のトランスジェニック植物は31日後に無病徴であった。23日後に、トランスジェニック植物の先端46cmを切り取り、新たな成長およびさらなるストレスを誘導した。若く、勢いよく成長している新しい苗芽は接種31日後にいずれの病徴も示さなかったが、接種45日後に各々33%、31%および45%のTSWV-BL、-T91および-Bが葉病徴または茎病徴を示していた。これらの結果は、あるトランスジェニック植物は耐性で、またあるものは感染に対して免であることを示している。
【0094】
かくして、本発明の1つの態様によれば、TSWV-BL単離株のNP遺伝子を発現するトランスジェニック植物は、同種のTSWV-BL単離株および同一血清群の異種単離株(アルカンサスおよび10W体菜)の感染に対して高い抵抗性を示した。さらに重要なことには、抵抗性は他の血清群からベゴニア単離株に至るまで有効である。略言すれば、TSWV-BLの核蛋白質遺伝子を発現するトランスジェニックタバコ植物はTSWVおよびINSVの双方に抵抗性を示し、その保護は遠縁のINSVに対する核蛋白質によって、ならびに同種および近縁のTSWV単離株に対する核蛋白質遺伝子リボヌクレオチド配列によって媒介されているようであることを前記のことは明らかに記載している。異なったTSWV単離株に対して広範囲スペクトル抵抗性の構築植物が示されたのはこれが初めてである。
【0095】
一般的に、外皮蛋白質保護は、免疫ではなくて、感染および病徴発現における遅延および/低下を示すが、本発明は、無病徴でかつ感染ウイルスフリーである有意に高いパーセンテージのトランスジェニック植物を提供する。温室条件下におけるこれらの植物の抵抗性はそれらの生涯を通して保持され、より重要なことは、前記したごとくそれらの子孫にも受け継がれた。
【0096】
たとえあるとしてもほとんどTSWV-BL NPを産生しないトランスジェニック植物が、同種の単離株、およびTSWVの同一血清群内の他の近縁の単離株による感染に高い抵抗性を示すことが本発明において認められた一方で、高レベルのNP遺伝子を発現しているものにおいては保護が認められなかった。
【0097】
TSWVの生物学的多様性は詳細に報告されており、トマトのごとき栽培植物においては遺伝的抵抗性を克服することが報告されている。かくして、多種のTSWV株に対して抵抗性を示すトランスジェニック植物を開発することが極めて重要である。本発明は、この抵抗性を付与するウイルスNP遺伝子を利用してそれが行われるであろうこと、ならびに多様なTSWV単離株にこの抵抗性が示されるであろうことを示している。かくして、TSWV NP遺伝子の発現が種々のTSWV単離株に対して高レベルの抵抗性を付与できるという本発明の知見は、膨大な量の商業的重要性を有する。
【0098】
他の研究のシリーズにおいて、プラスミドBIN19−N+を構築し、実施例IVに従ってアグロバクテリウム・チュメファシエンス(A.tumefaciens)LB4404株に移入し、実施例Vに従いニコチアナ・ベンサミアーナ(Nicotiana benthamiana)に移入した。INSV−Begおよび−LIのヌクレオキャプシド遺伝子をオリゴマープライマーINSV−A(5'−TACTTATCTAGAACCATGGACAAAGCAAAGATTACCAAGG)およびINSV−B(5'−TACAGTGGATCCATGGTTATTTCAAATAATTTATAAAAGCAC)で増幅し、INSV単離体のヌクレオキャプシド遺伝子の5'−コーディング領域および3'−非コーディング領域のそれぞれにハイブリダイズさせた。増幅したヌクレオキャプシド遺伝子断片を実施例IIIに従って精製し、実施例IVに従って消化して配列決定した。
【0099】
合計24のN+(pBIN19−N+で形質転換された)および18N−(ベクターpBIN19で形質転換された)形質転換エヌ・ベンサミアーナ植物を土壌に移し温室で発育させた。全てのN+系列は4−5の葉段階において遺伝子配列を含んでいることがPCRにより確認された。N蛋白質の蓄積の相対レベルはTSWV−BLN蛋白質の抗体を用い、各々独立したR0トランスジェニッククローン系においてDAS−ELISAによって評価した。24のN+系のうち、2つが0.50−1.00のOD405nmの値を有し、17が0.02−0.10の間、残りの5つが0.02より少なかった。健康なエヌ・ベンサミアーナまたはトランスジェニックN−植物は0.02−0/02のOD405nmの読みを与える。R0植物の全てが自家受粉され、以下のトランスジェニック系:(1)N−−2および−6、pBIN19ベクターのみを含む対照トランスジェニック系、(2)N+−28、N蛋白質の検出不可能な量を産生したトランスジェニック系(OD405nm=0.005);(3)N+−21、低レベルのN蛋白質を産生するトランスジェニック系(OD405nm=0.085);および(4)高レベルのN蛋白質を蓄積する2つのトランスジェニック系であるN+−34および−37(OD405nm=0.50−1.00)からの種子を植菌試験のためのカナマイシン選択培地(300μg/ml)上で発芽させた。次いで、これら6つの系をノーザンハイブリダイゼーションによって分析し;N遺伝子転写体の強度はELISA反応のレベルとよく相関していた。
【0100】
6つのR0系からのトランスジェニック実生はカナマイシン選択培地上で種子を発芽させることによって選択し、これらの実生を5種のトスポウイルスで接種した。接種されたR1植物は、もしウイルス病徴が、接種葉に観察されれば感受性と見なした。漏れの可能性を排除するために、トランスジェニック対照N−植物をトランスジェニックN+植物の各々の接種で常に用いた。加えて、各接種抽出物を、まずN+植物を接種し、次いで対照N−植物を接種するのに常に用いた。この一連の研究の結果を以下に示す:
【0101】
エヌ・ベンサミアーナ黄化壊疽ウイルス(TSWV)のヌクレオキャプシド
(N)蛋白質を発現するR1植物の、トスポウイルスでの接種に対する反応性
感染植物数/接種植物数b
TSWV単離株 INSV単離株
R0系 ELISAa BL 10W Beg LI TSWV-B
N--2/-6 <0.02 32/32 32/32 32/32 20/20 32/32
N+-28 0.005 16/16 16/16 15/16 16/16
N+-21 0.085 9/40 17/40 39/40 18/20 40/40
N+-34 0.715 25/28c 28/28 23/28c 28/28
N+-37 0.510 26/28c 22/22 21/28c 16/20c 22/22
a R1植物が由来するR0系のaELISAデータ;
b 感染したエヌ・ベンサミアーナ植物の30倍希釈葉抽出物を3−5葉段階の3枚の植物葉に適用した。各抽出物を常に用いてN+植物を接種した。データは少なくとも接種後2カ月毎日収集し、全身感染植物数/接種植物数として表した;cほとんど全ての感受性R1植物は病徴の出現の顕著な遅延を示すことを示す。
【0102】
上記の表で示したように、対照系N−−2および−6からの全てのR1植物は、試験した全てのウイルスの接種後5−8で全身的な病徴を示した。N+−28系からのR1植物のいずれも検出可能な量のN蛋白質を産生せず、INSV−Begを接種した1の植物を除き全てがこれらのウイルスに感受性であった。接種前に採取したこのN+−28R1植物のリーフディスクのELISA分析により、INSV−Beg抵抗性表現型を保有すると同定された植物が高レベルのN蛋白質を蓄積することが明白に示された(全ての他のN+−28R1植物に対しOD405nmが<0.02であるのと比較しOD405nm=0.78)。
【0103】
N遺伝子を低いレベルで発現しているN+−21系は同種(78%)および近縁のTSWV−10W(57%)単離株に対して良好な抵抗性を示し、は2個のINSV単離株に対しほとんど耐性を示さず(3%および10%);3のN+−21植物のみが、INSV単離株を接種した時に抵抗性の表現型を示した。これらのINSV−耐性N+21 R1植物からの葉の試料は非常に高いELISA反応を示し(OD405nm 0.5ないし1.00)、かくして感受性N+−21植物よりも多量のN蛋白質を与えた(OD405nm 0.02ないし0.50)。N遺伝子の発現が高い系はINSV単離株に対し最も高い抵抗性を示し(18%-5%)、次いで同種TSWV−BL単離株(7%および11%)であり、一方、植物のいずれもTSWL−10Wに対し抵抗性を示さず;しかしながら、INSVまたはTSWV−BLで感染されたN+−34および−37R1植物は病徴の発現において様々な長さの遅れを示す。これらの4つのトランスジェニックN+系からのR1植物のいずれもTSWV−Bに抵抗性でなかった;N+−34および−37系からのR1植物のいくつかは病徴の出現に少しの遅延を示した。
【0104】
N+R1植物におけるN蛋白質産生レベルが異なるトスポウイルスに対する抵抗性と関連しているか否かを決定する実験において、上記表中の接種N+R1植物は、元のR0植物とは関係なく接種前に採取された組織のそのELISA反応の強度に基づき4つの群に再び組織した。N蛋白質を低レベルで発現するN+R1植物(0.02−0.2OD)は、TSWV−BLと−10Wに対し高い抵抗性(100%および80%)を示すが、INSV−Begおよび−LIに対しては全て感受性であり、これは対照のN−植物に対する病徴出現において検出可能な遅延はないことを示す。対照的に、高いレベルのN蛋白質を有するほとんどすべてのN+R1植物(0.20−1.00OD)は、TSWV−BL、INSV−BegおよびLIに対して様々なレベルの保護を示し、病徴の出現の短い遅延から、これらの植物のほとんどが対照N−植物に対する病徴の発展における遅延の様々な長さを示す、完全な耐性の範囲にある。TSWL−10Wに対する高い発現因子において保護は観察されなかった。加えて、N遺伝子発現のレベルに拘らず、N+植物はTSWV−Bに対して抵抗性でなかった;しかしながら、高レベルでN蛋白質を産生するN+R1植物では、短い遅延の病徴の出現が観察された。全ての対照N−R1植物および検出不可能なELISA反応を起こす(0ないし0.02OD)トランスジェニックN+R1植物は試験したすべてのトスポウイルスに対し感受性であった。
【0105】
TSWV−BLヌクレオキャプシド遺伝子を発現するエヌ・ベンサミアーナプロトプラストにおいて遠縁INSVの複製阻害も実験した。これらの実験において、全てのINSV−LIウイルス粒子を用いて3つのトランスジェニック系から単離したプロトプラストを感染させて、形質転換の産物が、いかに入って来るウイルスに影響するかを調べた。ウイルスの複製は、INSV N蛋白質に特異的な抗体を用いてトランスジェニックプロトプラストにおいてINSVを感染するN蛋白質のレベルを測定することによって決定した。DAS−ELISA分析は、与えられた系からの全ての子孫が比較的均一であって、ほとんど全てのR1の子孫が、その親であるトランスジェニック系と類似したトランスジェニックN遺伝子の発現レベルを与えることを示した。これらの結果は、その親系のそれに基づくR1の集団の発現レベルの予測を可能にした。低い発現因子の系N+−21のR1植物由来のプロトプラストはINSV−LIの複製を支持するが、一方、高い発現因子系N+−37のR1植物からのプロトプラストは、低レベルのウイルスの複製が観察される接種の42時間後までこれを支持しなかった。様々な時間間隔(例えば0、19、30および42時間)で同じプロトプラストを、形質転換遺伝子の発現レベルを検査するために、TSWL−BL N蛋白質に特異的な抗体を用いたDAS−ELISA法によっても分析した。予測されるように、N+-21 R1植物からのプロトプラストは比較的低レベル(0.338-0.395,OD405nm)を産生するのに対し、N+−37 R1植物からのプロトプラストは高いレベルを蓄積した(0.822−0.865,OD405nm)。発現レベルは全ての時点で一定であることが判明した。
【0106】
本発明の本態様において、低量のTSWV−BL N蛋白質を蓄積するトランスジェニック・エヌ・ベンサミアーナ植物は同種および遠縁の(TSWV-10W)単離株に高度に対して抵抗性であるが、一方この蛋白質を多く蓄積する植物は同種および遠縁の(INSV−BegおよびINSV−LI)ウイルスの両方に対する適度に保護を有することが示された。より重要なことには、これらの知見は、トランスジェニック・エヌ・ベンサミアーナ植物(INSVの系統的な宿主)はINSV−BegおよびINSV−LI単離株に対し保護されることを示す。
【0107】
上記で示したように、本発明者らは、TSWVのN遺伝子を発現するトランスジェニック植物は、同種単離株に対して抵抗性であり、TSWV−BL遺伝子を発現するかかる植物はTSWVおよびINSVの双方に抵抗性であることを示した。同種および近縁の単離株に対する最良の抵抗性はN蛋白質を低レベルで蓄積しているトランスジェニック植物で見い出されるが、一方TSWV−BL N蛋白質の高レベルを有するトランスジェニック植物は血清学的に遠いINSV単離株により抵抗性であることも示された。この観察によって、本発明者らは、同種および近縁の単離株からの観察される保護における翻訳されたN蛋白質産物の役割に気付き、また植物ゲノムに挿入されたN遺伝子自体か、それともその転写物が保護に関与するかを憶測した。この仮説を試験するために、プロモーターを含まないN遺伝子を含むか、あるいはセンスまたはアンチセンスの翻訳不可能なNコーディング領域を発現するトランスジェニック植物を作出した。発見されたことは、センスおよびアンチセンスの翻訳不可能なN遺伝子RNAは同種および近縁の単離株に対する保護を供給すること、およびこれらのRNA媒介保護は、それぞれのRNA種を低いレベルで合成する植物において最も有効であって、ウイルス複製の阻害を通じて達成されるようである、ということである。
【0108】
より具体的には、トランスジェニック植物に導入されたコーディング配列を図7に示す。示されるように、pBIN19−N構築物は植物形質転換ベクターpBIN19に挿入された無プロモーターN遺伝子を含む(実施例IV参照)。全ての他の構築物は二重のCaMVの35Sプロモーター、アルファルファモザイクウイルスの5'−末端の翻訳リーダー配列およびノパリンシンターゼの3'−非翻訳/ポリアデニル化配列を含む。pBI525は植物発現ベクターであって、この研究においては対照として用いられ;pBI525−mNはN遺伝子の変異(翻訳不能)形を含み;pBI525−asNは翻訳不能N遺伝子のアンチセンス型を含む。変異体N遺伝子の5’末端における1個のヌクレオチドの欠失はダッシュの記号によって示される。ATGコドンに下線を施し、インフレームの終止コドンは太字で示す。
【0109】
実施例VIII
TSWL−BL N遺伝子のプライマー特異的突然変異誘発およびクローニングは以下のごとくに行った:
N遺伝子の全長は、ここに引用して本明細書の一部とみなすファイトパソロジー(Phytopathology)第82巻:1223頁(1992年)に記載されるごとく、逆転写およびポリメラーゼ鎖反応により得た。翻訳不能N−コーディング配列は、オリゴマープライマーA、(AGCATTGGATCCATGGTTAACACACTAAGCAAGCAC)(TSWV−BL N遺伝子の3’−非暗号領域におけるS RNAと同一)、およびB、(AGCTAATCTAGAACCATGGATGACTCACTAAGGAAAGCATTGTTGC)(N遺伝子の5'末端においてS RNAに相補的)を用いたRT−PCRにより同様に作られた。後者のオリゴマープライマーは転写開始コドンの直後のフレームシフト変異および起こり得る翻訳のリードスルーを阻止するためのいくつかの終止コドンを含んでいる。無傷のおよび変異のN遺伝子断片は実施例IIに記載したごとく1.2%アガロースゲルによって精製した。ゲル精製した無傷のおよび変異のN遺伝子断片を適当な制限酵素で消化し、それぞれ実施例IVに記載されているごとく、直接BamHI/およびXbaI消化の植物形質転換ベクターpBIN19およびNcoI消化の植物発現ベクターpBI525にクローンした。得られたプラスミドを同定し、カリフラワー・モザイク・ウイルス35Sプロモーターに対し、無傷の無プロモーターN遺伝子、およびセンスおよびアンチセンスの向きで変異コーディング配列を有するpB1525−mNおよびpB1525−asNを含むpBIN19−Nと命名された。ニコチアナ・タバカム(Nicotiana tabacum)プロトプラストにおける一過性発現分析により発現カセット中の変異Nコーディング配列の翻訳可能性をチェックし;センスまたはアンチセンスの変異Nコーディング配列を含む発現カセットをHindIII/EcoRIによる部分消化(なぜならNコーディング配列は内部のHindIIIおよびExoRI配列を含むので)により切り出し、同酵素で既に切断されている植物形質転換ベクターpBIN19に連結した。pBIN19-Nと同様得られたベクターを、実施例IVに記載した方法を用いてエイ・チュメファシエンスLBA4404株に移入した。エヌ・タバカム・バール・ハバナ・シイブイ(N.tabacum var Havana cv 423)のリーフディスクを、様々な構築物を含有するエイ・チュメファシエンスのLBA4404株で接種し、得られたトランスジェニック植物を自家授粉させ、種子を選択的カナマイシン培地上で発芽させた。
【0110】
PCRは上記のごとく各R0トランスジェニック系で行った。オリゴマープライマーAおよびBを用いてTSWL−BLのN−コーディング配列の存在を決定した。オリゴマープライマー35S−プロモーター(CCCACTATCCTTCGCAAGACCC)をオリゴマープライマーAもしくはBのいずれかと組み合わせて、植物ゲノムに挿入された変異体のNコーディング配列の(CaMV 35Sプロモーターに対する)向きを確認した。DAS−ELISA法を用い、TSWL−BLN蛋白質に対するポリクローナル抗体を用いてトランスジェニック植物においてN蛋白質を検出した。ノーザンブロットによるトランスジェニック植物におけるRNA転写レベルを見積もるために、全植物RNAをナポリ(Napoli)の方法に従って単離し[ザ・プラント・セル(the Plant Cell)第2巻:279頁(1990年)参照]、ホルムアルデヒド含有アガロースゲルで分離した(10μg/レーン)。次いで、該アガロースゲルをエチジウムブロミドで染色して各レーンの全植物RNAの均一性を確認した。ハイブリダイゼーションの条件は製造者によるジーン・スクリーン・プラス(GeneScreen Plus)のプロトコールに記載されている通りである。得られたシグナルブロットを各ブロットに含まれる対照レーンのN遺伝子転写物バンド(高レベルのN遺伝子を産生するmNR1植物)に基づき比較し標準化した。100および150の間の密度値(ヒューレット・スカンジェット・アンド・イメージ・アナリシス・プログラム(Hewlet ScanJet and Image Analysis Program))を示したトランスジェニック植物は高い発現因子と見積もり、一方15および50の間の密度値を有するものは低い発現因子と見積もった。
【0111】
トスポウイルスでのトランスジェニック植物の接種は、特記する以外は前記したごとく行い、接種は3−4葉段階において行った。
タバコ・プロトプラストはR1植物由来の表面を殺菌した葉から調製した
[ツェット・プフランツェフィジオル(Z.Pflanzanphysiol)第78巻:453頁(1992年)に変更を加えたので、参照のこと]単離されたプロトプラスト(6×106プロトプラスト)をPEG法[プラント・アンド・モレキュラー・バイオロジー(Plant Mol.Biol.)第8巻:363頁(1987年)参照]を用いて、OD260nm値0.68の精製したTSWL−BLウイルス粒子抽出物で形質転換した。次いで、形質転換したプロトプラストを暗所にて26℃で終密度1×106プロトプラスト/ml培養培地になるまで培養した。様々な間隔のインキュベーションの後で、培養したプロトプラストをW5溶液で2度洗浄しエンザイム結合緩衝液中、浸透圧のショックにより溶解させた。ウイルスの増殖(複製)はDAS−ELISA法を用いてウイルスのN蛋白質を測定することによって見積もった。
【0112】
記載したごとく、本発明の一の態様は、全く検出できないか僅かしか検出できない量のN蛋白質を生産するトランスジェニック・タバコは同種または近縁の単離株に対し抵抗性であることを示す。この結果は、観察された抵抗性が、入ってきたウイルスN遺伝子と、トランスジェニック植物で産生されたN遺伝子転写物またはNコーディング配列それ自体のいずれかのトランス相互作用によるものであることを示唆する。核N遺伝子の存在が役割を果たすかどうかを試験するために、2つのP0N系からのトランスジェニックP0N R0系およびR1植物を4種のトスポウイルス(TSWV−BL、TSWV−10W、INSV−BegおよびTSWV−B)で攻撃した。無病徴性の植物は抵抗性と評価されたのに対し、何らかの病候を示す植物は感受性であると評価した。全ての接種したR0およびR1植物はウイルスに感受性であった。
【0113】
さらにNトランス遺伝子(transgene)の転写物が保護に関与する可能性を試験するために、センスまたはアンチセンスN遺伝子転写物いずれかを産生するが、N蛋白質は産生しない多数のR0トランスジェニック植物に同種単離株で接種した。結果を以下の表に示す:
【0114】
【表2】
【0115】
接種後7ないし9日で典型的な全身的病徴を示す対照とは異なり、21のmN植物のうち16、および8のasNのうち5のがそのライフサイクルの間中ずっと無病徴であった。接種前に採取された葉の組織のノーザンブロット分析は、全ての抵抗性R0系が低レベルのセンスまたはアンチセンスのN遺伝子RNAを産生するが、一方感受性R0系はRNA種を産生しないかまたは高レベルで産生する。このデータは、トランスジェニック植物のTSWV−BLに対する抵抗性はN遺伝子転写物のその相対的なレベルに関連していることを示唆したので、高いまたは低いN遺伝子転写レベルの4種のmNおよび3種のasN R0系からのトランスジェニックの子孫をカナマイシン含有培地上で発芽させることによって選択した。これらのトランスジェニック植物は、2種のasN系からのいくつかのR1植物は6ないし7葉段階で接種した以外は、3ないし4葉段階において4種のトスポウイルスに対する抵抗性を試験した。結果を以下の表にまとめる:
【0116】
【表3】
【0117】
高い発現因子系mN−2およびmN−7からの全てのR1植物は試験した全てのトスポウイルスによる感染に対して感受性であり、これらの植物は対照と比較して病徴の遅延を示さなかった。対照的に、低い発現因子系mN−13および−18からのR1植物の高い割合のものが同種(TSWV−BL)および近縁(TSWV−10W)単離株に対して抵抗性であったが、遠縁トスポウイルス(INSV−BegおよびTSWV−B)による感染に対しては抵抗性がない。低い発現因子のR0系からのasN R1植物の抵抗性は接種に用いたTSWV単離株によって顕著に影響された。低い発現因子系asN-1および-9からの小さいR1植物(3-4葉段階)の1つを除いて全てが感染したが、同種TSWV−BLまたは近縁TSWV−10Wの単離株を接種した場合は病徴の出現の遅延があった。対照的に、系asN−9からの大きいR1植物(6−7葉段階)のほとんどが両方の単離株に対して抵抗性であった。比較すると、対照R1植物およびasN−4のごとき高発現因子系からのR1植物は、植物の大きさによらずいずれの単離株に対しても抵抗性を示さなかった。アンチセンスRNA媒介される保護は遠縁のINSV−BegおよびTSWV−B単離株による感染に対して有効ではなかった。
【0118】
上記の2つの表に示されたデータの分析によって、センスおよびアンチセンスのRNA−媒介保護はN遺伝子の低い発現因子においてのみ観察されることが示された。高レベルのアンチセンスN遺伝子転写物を産生するR1 asN植物は対照の植物と同程度に感受性があった。対照的に、asNの低い発現因子は、3−4葉段階で接種した場合は病徴の出現に遅延を示し、6−7葉段階で接種した場合はレベルの増加を示した。
【0119】
翻訳不可能Nコーディング配列のセンスまたはアンチセンス型を発現しているタバコ・プロトプラストのウイルス複製の阻害も認められた。この場合には、TSWV−BLの全ウイルス粒子調製物を、トランスジェニック系から単離したプロトプラストをトランスフェクトするのに用いて、入って来るウイルスの複製に与えるセンスまたはアンチセンスのN遺伝子転写物の影響を調べた。ウイルス複製を、トランスフェクトしたプロトプラストに入って来るウイルスのN蛋白質のレベルを測定することによって決定し、高レベルの各RNA転写物を産する植物(mN−7およびasN−4)からのプロトプラストはウイルスの複製を支持し、一方mNの低い発現因子(mN−18)からのプロトプラストは支持しないことが判明した。asNの低い発現因子(asN−9)からのプロトプラストははるかに低いレベルのウイルス複製を支持した。
【0120】
従って、本発明の本態様において、本発明者らは、翻訳不可能N遺伝子コーディング配列のセンスまたはアンチセンス型を発現するトランスジェニック植物は、同種(TSWV−BL)または近縁(TSWV−10W)トスポウイルスには抵抗性であるが、遠縁の(INSV−BegまたはTSWV−B)トスポウイルスには抵抗性でないことを示した。以下の表は、様々な形態のTSWV-BL N遺伝子を発現するトランスジェニック・タバコ間のトスポウイルスに対する抵抗性の比較を示す:
【0121】
【表4】
【0122】
これらの結果により、TSWVでのRNA媒介保護のための本発明の前に述べた態様が確認され、拡大される。さらに、該保護は高レベルのN遺伝子転写物を生産する植物においてよりも、低レベルのN遺伝子転写物を生産する植物において観察され、本明細書中で報告する初期の研究は、高レベルのTSWV−BL N蛋白質を産生するタバコ植物がINSV−Begに対する抵抗を呈することを示すにも拘わらず、この付加的なデータは、INSV−Begに対する抵抗性がセンスおよびアンチセンス型の翻訳不能N遺伝子を発現するトランスジェニック植物では観察されないことを示すので、かくしてINSV−BEGに対する保護はN蛋白質の存在によるものであって、N遺伝子転写物によるものではないことを明白に示唆することを示す。従って、2の異なる機構が、本発明によるTSWVおよびINSVトスポウイルスに対する保護に関与しているように思われる。一の機構はN遺伝子転写物を含み(RNA−媒介)、もう一個はN蛋白質を含む(蛋白質媒介)。加えて、プロトプラスト実験の結果は、N遺伝子のRNA−媒介保護は、ウイルス複製を阻害する過程を通じて達成されることと、上記の表に含まれるデータは、遠縁のINSV−Beg単離株に対する保護はTSWV−BIのN蛋白質によって授与されるのであって、遺伝子転写物によって授与されるのではないことを示唆する。
【0123】
最後に、本発明のもう一個の態様−すなわちトスポウイルス核蛋白質遺伝子の一部はトスポウイルスによる感染に対するトランスジェニック植物の保護を提供するためにさらに研究を行った。N遺伝子RNAは同種および近縁TSWV単離株に対して保護し、一方で、N蛋白質は同種単離株および遠縁のINSV単離株に対し保護すること;N遺伝子のRNE媒介保護は、N遺伝子を低いレベルで発現している植物で有効であるが、一方N蛋白質媒介保護はN蛋白質の高いレベルの蓄積を要すること;およびN遺伝子のRNA媒介保護は、ウイルス複製の阻害を通じて達成されることは上記ですでに示した。この以前のデータに基づき、本発明者らは次に、N遺伝子の一部がウイルスによる感染に対して働くかどうかを決定することに着手した。本発明らは、下記に議論するように、N遺伝子配列の約半分を発現しているトランスジェニック植物はウイルスに対して抵抗性であることを見い出した。
【0124】
以下に、本発明のこの最後の態様を示すためにTSWV−BLの半分のN遺伝子断片のクローニングを記載する。翻訳可能なまたは翻訳不可能なN遺伝子の両方の、第一および第二の半分を、上記に記載したように逆転写、および次いでのPCRによって作成した。図8に示すように、翻訳不可能なN遺伝子断片の半分の5’末端でのヌクレオチドの欠失または挿入はダッシュの記号によって示し;ATGコドンは下線を施し、翻訳不可能なN遺伝子の半分の断片の開始コドンのすぐ後にある可能な終止コドンは太字で示す。
【0125】
N遺伝子の第一の半分はオリゴプライマーi(5'−TACAGTGGATCCATGGTTAAGGTAATCCATAGGCTTGAC)(これはTSWV−BLN遺伝子の中央領域に相補的)および翻訳可能な第一の半分のN遺伝子断片用のii(5'−AGCTAACCATGGTTAAGCTCACTAAGGAAAGCATTGTTGC)、もしくはiii翻訳不可能な第一の半分のN遺伝子断片のための(5'−AGCTAATCTAGAACCATGGATGACTCACTAAGGAAAGCATTGTTGC)を用いてRT−PCRによって生産され、後の方の2つのオリゴマープライマーはN遺伝子の5'末端と同一である。同様に、N遺伝子の第二の半分は、オリゴマープライマーiv(5'−AGCATTGGATCCATGGTTAACACACTAAGCAAGCAC)(これはTSWV−BLN遺伝子の3'非コード領域に相補的)および翻訳可能な半分のN遺伝子断片のためのv(5'−TACAGTTCTAGAACCATGGATGATGCAAAGTCTGTGAGG)、または翻訳不可能な半分のN遺伝子断片のためのvi(5'−AGATTCTCTAGACCATGGTGACTTGATGAGCAAAGTCTGTGAGGCTTGC)を用いてRT−PCRによって生産し、後の方の2個のオリゴマープライマーはN遺伝子の中央領域と同一である。オリゴマープライマーiiiは翻訳コドンのすぐ後にフレームシフト変異および、可能な翻訳のリードスルーを阻害するためのいくつかのインフレームの終止コドンを含む。
【0126】
半分の遺伝子断片を上記のごとく1.2%アガロースゲルで精製し、ゲルで単離した断片を制限酵素NcoIで消化し、NcoI消化した植物発現ベクターpBI525にクローンした。得られたプラスミドを同定し、(1)第一の半分の翻訳可能なN遺伝子を含むpBI525−1N、(2)第一の半分の翻訳不可能なN遺伝子を含むpBI525−1N'、(3)アンチセンスの向きで第一の半分の翻訳可能なN遺伝子を含むpBI525−1N−、(4)第二の半分の翻訳可能なN遺伝子を含むpBI525−2N、(5)第二の半分の翻訳不可能なN遺伝子を含むpBI525−2n'、(6)アンチセンス向きでの第二の半分の翻訳可能なN遺伝子を含むpBI525−2N−と命名した。次いで、発現カセットをHindIII/EcoRIで消化することによってプラスミドpBI525から切り出し、上記のごとく、同じ酵素ですでに切断した植物形質転換ベクターpBIN19に連結した。プラスミドpBIN19と同様得られたベクターを、ホルスターズ(Holsters)(前掲)によって記載された方法を用いてエイ・チュメファシエンス(A.tumefaciens)に移入した。エヌ・ベンサミアーナ(N.benthamiana)のリーフディスクに、様々な構築物を含むエイ・チュメファシエンスのLB4404株を接種した。トランスジェニック植物を自家授粉させ、種子を上記のごとくカナマイシン上で発芽させた。
【0127】
PCRおよびノーザンハイブリダイゼーションPCRによるトランスジェニック植物の分析を、以前記載されたごとく各R0トランスジェニック系において行った。オリゴマープライマーiないしviを、TXWV−BLのNコーディング配列の存在を決定するために用いた。オリゴマープライマー35S−プロモーター(実施例VIII照)を上記のオリゴマープライマーの一つと組み合わせて、植物ゲノムに挿入された半分の遺伝子配列の向き(CaMV35Sプロモーターに対する)を確かめた。ノーザン分析は、実施例VIIIに記載されるごとくに行った。
【0128】
TSWV(TSWV−BL)のレタス単離株を用いてトランスジェニック植物を攻撃した。上記のごとく3−4葉段階のテスト植物を用いて接種した。逃避の可能性を防ぐために、対照の植物を各実験において用い、各接種抽出物を用いてが、まず第一にトランスジェニック植物、次いで対照植物に接種した。
【0129】
本発明の本態様におけるさまざまな構築物を図8に示す。翻訳可能および翻訳不可能な半分のN遺伝子断片をRT−PCRにより合成し、次いで植物発現ベクターpBI525に直接クローンした。RT−PCRによる翻訳不可能な半分のN遺伝子断片の産生のために用いたオリゴマープライマーiiiおよびviは、翻訳開始コドンのすぐ後に変異を有し、得られたリーディングフレームは可能な翻訳のリードスルーを阻害するためのいくつかの終止コドンを含む。従って、第一および第二の半分の翻訳不可能なN遺伝子断片は共に、植物に導入された際に、端を切り取ったN蛋白質断片を産生するのは不可能であるに違いない。次いで、翻訳可能なおよび翻訳不可能な半分のN遺伝子断片を共にセンスまたはアンチセンスの向きでベクターpBI525のCaMV35Sプロモーターの下流に入れた。TSWV−BLの半分のNコーディング配列の発現は、従って、発現ベクターpBI525のアルファルファモザイクウイルス(ALMV)の翻訳されないリーダー配列に融合した二重のCaMV 35Sプロモーターによって制御された。積み重ねられた二重のCaMV 35Sプロモーターを用いる発現ベクターは、一個の35Sプロモーター配列を有する類似のベクターよりも高レベルのmRNA転写を起こすことが公知である。発現カセットをベクターpBI525からパント(pant)形質転換ベクターpBIN19に移入した。対照のプラスミドpBIN19と同様に、得られたプラスミドを次いでエイ・チュメファシエンスLB4404株に移入した。トランスジェニック植物を図8に示すトランスジェニック系の命名と共に得た。
【0130】
全てのカナマイシン抵抗性トランスジェニック系はPCRによって、期待される向きで適切なNコーディング配列を含むことを確認した。種子用に成育させた各トランスジェニックR0系をノーザンブロットにより分析した。6つの1Nのうち6、6つの1N'のうち4、6つの1N−のうち6、6つの2Nのうち6、8つの2N'のうち7、および 7つの2N−トランスジェニックR0のうち6は、半分のN遺伝子RNAを産生することが判明した。
【0131】
トランスジェニックR0植物の一組を同種単離株TSWV−BLで攻撃した。無病徴の植物を抵抗性である評価し、一方でなんらかの病徴(局部の障害または全身的な感染)を示した植物を感受性であると評価した。全ての接種されたR0対照植物はウイルスに感受性であった;対照的に、9つの1N'のうち2、6つの1N−のうち2、10つの2N’のうち4、および8つの2N−R0系のうち一つがウイルス感染に完全に抵抗性であった。1Nおよび2N R0系のどちらとも高レベルの抵抗性を示したが、これらの植物のいくらかは病徴の出現に顕著な遅延を示した。
【0132】
トランスジェニックR0系のもう一組を種子の産生のために成熟させた。実生はカナマイシン含有培地で発芽させ、TSWV−BLを接種した。以下の表に示すように、対照の実生およびいくつかのトランスジェニック系からの実生は単離株に感受性であるのに対し、1N-151、1N'-123、および2N'-134系からの実生は、病徴の発現の遅延から完全な抵抗性まで変化するさまざまなレベルの保護を示した。
【0133】
感染させた植物の数/接種した植物の数
R0系 6DPI 15DPI 30DPI
対照 50/50
1N −149 17/17
1N −151 2/20 13/20 17/20
1N'−123 16/20 17/20 17/20
1N'−124 20/20
1N'−126 19/19
1N−−130 12/15 15/15
1N−−132 18/19 19/19
2N −155 20/20
2N'−134 0/20 10/20 10/20
2N'−135 19/19
2N−−142 20/20
2N−−143 20/20
上記の表において、感染したエヌ・ベンサミアーナ(N.benthamiana)からの
30倍希釈の抽出物を、3−4葉段階のトランスジェニック植物、次いで対照トランスジェニック植物に接種するのに用いた。DPI=接種後の日数
【0134】
要約として、本発明の本態様は、翻訳可能なまたは翻訳不可能なN遺伝子断片の第一のまたは第二のいずれかの半分を発現しているトランスジェニック植物は、同種TSWV−BL単離株に高度に抵抗性であることを示す。この結果は、N遺伝子の一部だけでウイルスに対する抵抗性に十分であることを示す。
【0135】
本発明の上記記載のすべての核酸およびアミノ酸配列のリストは以下の通りである:
【0136】
【0137】
従って、本発明者らは本発明の好ましい具体例を例示し記載したが、本発明の変化および修飾が可能であることが理解されるべきであり、本発明者らはそれゆえ発表された正確な用語を限定することを望むのではなく本発明に様々な用途および状態を付与するためになされる変化および交替を我々に役立たせることを望むものである。このような変化および修飾は、例えば、構造的に類似した核酸配列の置換を含むであろうが、ここにおいて、示される配列および変化した配列間の相違は、もしその変化した配列、即ち、上記記載のものと実質的に類似の結果をうむ配列にほとんど利点をもたらさない。従って、具体的に上記したそれらの配列の機能を実質的に変更することはない塩基(塩基配列中の)またはアミノ酸(ペプチド配列中の)の置換、欠失、挿入あるいは付加は本発明の態様の中に含まれると思われる。加えて、本発明が、本発明に従い植物へのトスポウイルスの感染に対する耐性または免疫を供給する様々な単離株のN遺伝子を一個のカセットにつなぐように修飾されること、およびトスポウイルスに対する広い抵抗性、特にTSWV−BL、TSWV−BおよびINSVに対する抵抗性を供給するためのトランス遺伝子(transgene)としてこのカセットを用いることは本発明者らの努力である。従って、このような変化および交替は本発明と等価である十分な範囲内で適切に意図され、以下の請求の範囲内にある。
その分野、またはそれが最も密接に関連した分野に属するいかなる当業者にも、同じ事をさせ使用させることができるように、十分で、明瞭で、完結で、正確な術語を用いて、本発明およびそこにおいてなされた方法および過程を記載した。
【図面の簡単な説明】
【0138】
【図1】図1は、本発明によるウイルスRNAからNP遺伝子をクローニングするための戦略を示す図である。
【図2】図2は、タバコ・プロトプラストにおける本発明によるトマト黄化壊疽病ウイルスのヌクレオキャプシド蛋白質(NP)遺伝子のイン・ビボ(in vivo)経時的発現を示す図である。
【図3】図3は、本発明によるTSWV-B S RNAにおける、配列決定したcDNAクローンの位置を示す図である。
【図4】図4は、本発明によるTSWV単離株の中での関係を示す樹系図である。
【図5】図5は、本明細書に記載したTSWV単離株の血清学的な関係を示す図である。
【図6】図6は、トランスジェニック植物におけるヌクレオキャプシド蛋白質(NP)の蓄積のレベルとTSWV単離株に対する抵抗性の度合との相関を示す図である。
【図7】図7は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BL N暗号配列を示す図である。
【図8】図8は、本発明の1つの態様によりトランスジェニック植物に導入したTSWV-BLの半分のN遺伝子断片を示す図である。
【特許請求の範囲】
【請求項1】
配列番号:15の核酸分子;
配列番号:19の核酸分子;および
配列番号:13のアミノ酸配列をコードする核酸分子
よりなる群から選択される単離された核酸分子。
【請求項2】
核酸分子が配列番号:19のヌクレオチド配列を含む請求項1記載の単離された核酸分子。
【請求項3】
核酸分子が配列番号:13のアミノ酸配列をコードする請求項1記載の単離された核酸分子。
【請求項4】
核酸分子が配列番号:15のヌクレオチド配列を含む請求項1記載の単離された核酸分子。
【請求項5】
請求項1記載の核酸分子によってコードされた単離された蛋白質またはポリペプチド。
【請求項6】
蛋白質またはポリペプチドが配列番号:13のアミノ酸配列を含む請求項5記載の単離された蛋白質またはポリペプチド。
【請求項7】
(i)トスポウイルスTSWV−Bの全長ヌクレオキャプシド蛋白質または(ii)トスポウイルスTSWV−Bの全長ヌクレオキャプシド蛋白質の長さの少なくとも約半分である全長未満の長さのヌクレオキャプシドポリペプチド断片のいずれかをコードする翻訳可能または翻訳不可能のいずれかのmRNAをコードする請求項3記載のDNA分子;
そのDNA分子の転写を可能とするDNA分子の5’側に操作可能に連結したプロモーター;および
DNA分子の3’側の終止配列;
を含むDNA構築体。
【請求項8】
DNA分子が配列番号:19のDNA分子からのものである請求項7記載のDNA構築体。
【請求項9】
請求項7記載のDNA構築体を含む組換えDNA発現系。
【請求項10】
請求項7記載のDNA構築体で形質転換された植物細胞。
【請求項11】
DNA分子が配列番号:19のDNA分子からのものである請求項10記載の植物細胞。
【請求項12】
請求項7記載のDNA構築体を含むトランスジェニック植物。
【請求項13】
DNA分子が配列番号:19のDNA分子からのものである請求項12記載のトランスジェニック植物。
【請求項14】
トランスジェニック植物がTSWV−Bに対して抵抗性である請求項12記載のトランスジェニック植物。
【請求項15】
請求項7記載のDNA構築体で植物細胞を形質転換し、次いで、形質転換された植物細胞から植物体を再生することを含み、
ここに、DNA分子が該トスポウイルスに対する抵抗性を植物体に付与することを特徴とするトスポウイルス抵抗性トランスジェニック植物の生産方法。
【請求項16】
DNA分子が配列番号:19のDNA分子からのものである請求項15記載の方法。
【請求項17】
植物がトスポウイルスTSWV−Bに対して抵抗性である請求項15記載の方法。
【請求項1】
配列番号:15の核酸分子;
配列番号:19の核酸分子;および
配列番号:13のアミノ酸配列をコードする核酸分子
よりなる群から選択される単離された核酸分子。
【請求項2】
核酸分子が配列番号:19のヌクレオチド配列を含む請求項1記載の単離された核酸分子。
【請求項3】
核酸分子が配列番号:13のアミノ酸配列をコードする請求項1記載の単離された核酸分子。
【請求項4】
核酸分子が配列番号:15のヌクレオチド配列を含む請求項1記載の単離された核酸分子。
【請求項5】
請求項1記載の核酸分子によってコードされた単離された蛋白質またはポリペプチド。
【請求項6】
蛋白質またはポリペプチドが配列番号:13のアミノ酸配列を含む請求項5記載の単離された蛋白質またはポリペプチド。
【請求項7】
(i)トスポウイルスTSWV−Bの全長ヌクレオキャプシド蛋白質または(ii)トスポウイルスTSWV−Bの全長ヌクレオキャプシド蛋白質の長さの少なくとも約半分である全長未満の長さのヌクレオキャプシドポリペプチド断片のいずれかをコードする翻訳可能または翻訳不可能のいずれかのmRNAをコードする請求項3記載のDNA分子;
そのDNA分子の転写を可能とするDNA分子の5’側に操作可能に連結したプロモーター;および
DNA分子の3’側の終止配列;
を含むDNA構築体。
【請求項8】
DNA分子が配列番号:19のDNA分子からのものである請求項7記載のDNA構築体。
【請求項9】
請求項7記載のDNA構築体を含む組換えDNA発現系。
【請求項10】
請求項7記載のDNA構築体で形質転換された植物細胞。
【請求項11】
DNA分子が配列番号:19のDNA分子からのものである請求項10記載の植物細胞。
【請求項12】
請求項7記載のDNA構築体を含むトランスジェニック植物。
【請求項13】
DNA分子が配列番号:19のDNA分子からのものである請求項12記載のトランスジェニック植物。
【請求項14】
トランスジェニック植物がTSWV−Bに対して抵抗性である請求項12記載のトランスジェニック植物。
【請求項15】
請求項7記載のDNA構築体で植物細胞を形質転換し、次いで、形質転換された植物細胞から植物体を再生することを含み、
ここに、DNA分子が該トスポウイルスに対する抵抗性を植物体に付与することを特徴とするトスポウイルス抵抗性トランスジェニック植物の生産方法。
【請求項16】
DNA分子が配列番号:19のDNA分子からのものである請求項15記載の方法。
【請求項17】
植物がトスポウイルスTSWV−Bに対して抵抗性である請求項15記載の方法。
【図1】

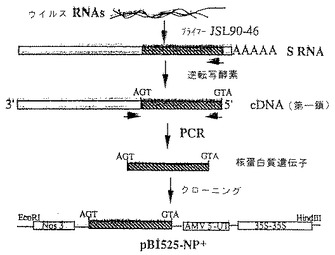
【図2】
【図3】
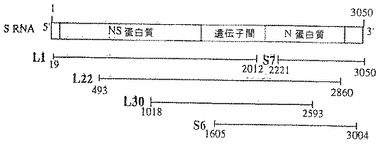
【図4】
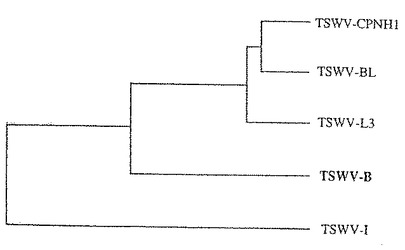
【図5】
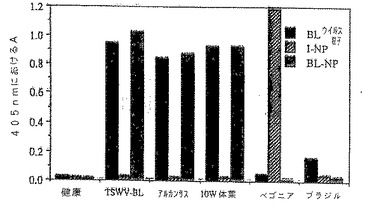
【図6】
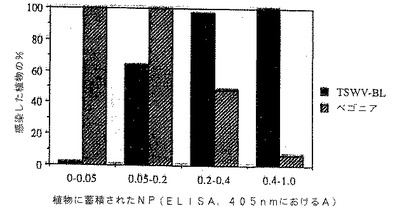
【図7】
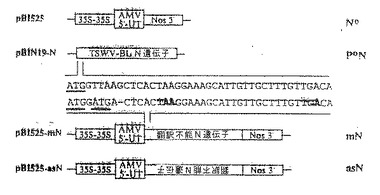
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2007−20579(P2007−20579A)
【公開日】平成19年2月1日(2007.2.1)
【国際特許分類】
【出願番号】特願2006−261120(P2006−261120)
【出願日】平成18年9月26日(2006.9.26)
【分割の表示】特願平6−517360の分割
【原出願日】平成6年1月27日(1994.1.27)
【出願人】(592035453)コーネル・リサーチ・ファンデーション・インコーポレイテッド (39)
【氏名又は名称原語表記】CORNELL RESEARCH FOUNDATION, INCORPORATED
【Fターム(参考)】
【公開日】平成19年2月1日(2007.2.1)
【国際特許分類】
【出願日】平成18年9月26日(2006.9.26)
【分割の表示】特願平6−517360の分割
【原出願日】平成6年1月27日(1994.1.27)
【出願人】(592035453)コーネル・リサーチ・ファンデーション・インコーポレイテッド (39)
【氏名又は名称原語表記】CORNELL RESEARCH FOUNDATION, INCORPORATED
【Fターム(参考)】
[ Back to top ]