
リシノール酸産生酵母の培養方法
【課題】リシノール酸産生酵母を、増殖性とリシノール産生性のバランスをとりながら効率よく培養する方法、およびリシノール酸の製造方法を提供する。
【解決手段】FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、を有することを特徴とするリシノール酸産生酵母の培養方法、並びに、該培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法である。
【解決手段】FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、を有することを特徴とするリシノール酸産生酵母の培養方法、並びに、該培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、酵母によりリシノール酸を産生させる方法に関する。より詳細には、FAH12遺伝子が導入された酵母の形質転換体を培養することにより、該形質転換体にリシノール酸を産生させる方法に関する。
【背景技術】
【0002】
現在、ウレタン製品等の化成品の原料は、専ら石油資源に頼っている。地球温暖化対策により、石油資源に代わる代替材料が模索されており、リシノール酸が注目されている。リシノール酸は、トウゴマ(Ricinus communis)を絞ったひまし油の80〜90%を占めており、ひまし油から精製したものが使用されている。しかし、トウゴマにはリシンという猛毒の蛋白質が含まれており、取り扱いに注意を要する上、植物資源であるため、安定供給には不安がある。そこで、ひまし油以外からリシノール酸を安全かつ安定して製造する方法の開発が望まれている。
【0003】
リシノール酸は、鎖長が炭素数18であり、不飽和結合が1つと12位に水酸基を持つ脂肪酸であり、Δ12−ヒドロキシラーゼにより、オレイン酸の12位に水酸基を導入することによって生産できる。野生型の酵母はΔ12−ヒドロキシラーゼを有していないため、リシノール酸を産生することはできない。そこで、本発明者らは、分裂酵母シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe、以下、S.ポンベ)が、脂肪酸組成としてオレイン酸の含有量が8割程度と高いことに着目して、S.ポンベに異種生物由来のΔ12−ヒドロキシラーゼ遺伝子を導入し、リシノール酸の生産を試みた。この結果、S.ポンベの形質転換体においてリシノール酸が生産されることが確認されたが、リシノール酸の生産量が多くなるほど宿主の増殖が遅くなることも分かった(非特許文献1参照。)。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】石川大輔、他8名、日本分子生物学会年会講演要旨集、2009年、第32巻、第2号、第70ページ。
【発明の概要】
【発明が解決しようとする課題】
【0005】
そこで本発明の目的は、リシノール酸産生酵母を、増殖性とリシノール産生性のバランスをとりながら効率よく培養する方法、およびリシノール酸の製造方法を提供することにある。
【課題を解決するための手段】
【0006】
本発明者等は、前記課題を解決すべく鋭意検討した結果、FAH12(Fatty acid hydroxylase)遺伝子が導入された酵母の形質転換体を、比較的高温で培養することにより、増殖阻害を抑制し得ること、また、高温培養により充分に培養させた後、温度シフトし、より低温で培養することによって、該形質転換体内のリシノール酸含有量を高められることを見出し、本発明を完成するに至った。
【0007】
本発明のリシノール酸産生酵母の培養方法、およびリシノール酸の製造方法は下記[1]〜[6]である。
[1]FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、
前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、
を有することを特徴とするリシノール酸産生酵母の培養方法。
[2]前記低温培養工程における前記形質転換体の培養温度が、10〜33℃である前記[1]のリシノール酸産生酵母の培養方法。
[3]前記酵母がシゾサッカロミセス(Schizosaccharomyces)属酵母である前記[1]または[2]のリシノール酸産生酵母の培養方法。
[4]前記FAH12遺伝子が、トウゴマ(Ricinus communis)、レスクェレラ・フェンドレリ(Lesquerella fendleri)または麦角菌(Claviceps purpurea)由来である前記[1]〜[3]のいずれのリシノール酸産生酵母の培養方法。
[5]前記[1]〜[4]のいずれのリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法。
【発明の効果】
【0008】
本発明のリシノール酸産生酵母の培養方法により、リシノール酸産生酵母を、増殖性とリシノール酸生産性のバランスをとりながら効率よく培養できる。
また、該形質転換体を用いた本発明のリシノール酸の製造方法により、産生されたリシノール酸を簡便に取得できる。
【図面の簡単な説明】
【0009】
【図1】pSL10−CpFAH12およびpSL10−LfFAH12のベクターの構築手順を模式的に示した図である。
【図2】pREP1−RcFAH12、pREP1−LfFAH12、およびpREP1−CpFAH12のベクターの構築手順を模式的に示した図である。
【図3】YEp352−CpFAH12のベクターの構築手順を模式的に示した図である。
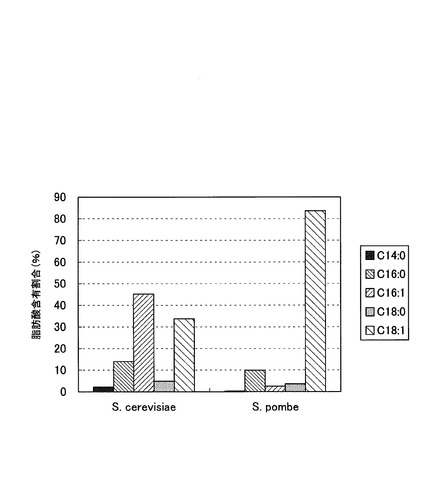
【図4】参考例1において、S.ポンベおよびS.セレビシエの脂肪酸含有量を測定した結果を示した図である。
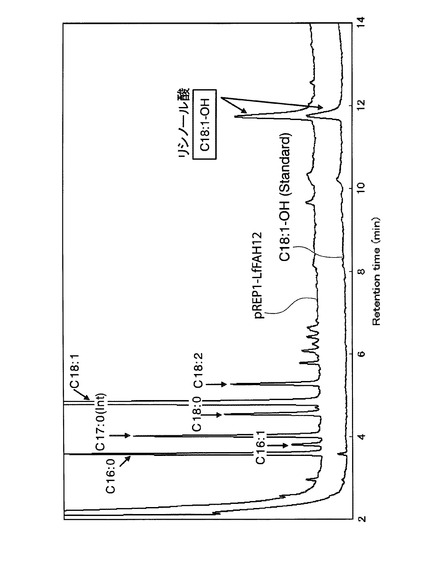
【図5】参考例3において、pREP1−LfFAH12を導入したS.ポンベの形質転換体のガスクロマトグラフ分析の結果のチャート図である。
【図6】実施例1において、各形質転換体をスポットしたプレートをインキュベートした後の各プレートの写真図である。
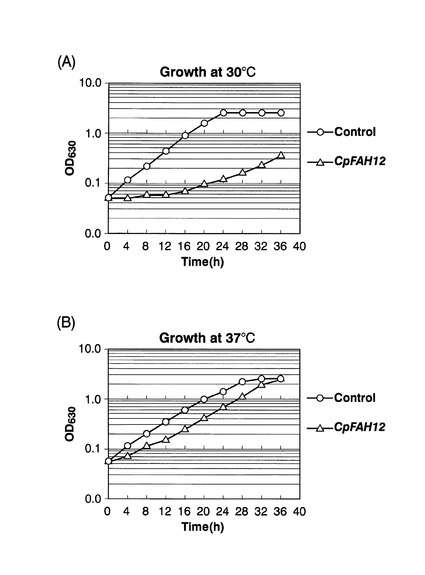
【図7】実施例2において、各形質転換体を、30℃または37℃で培養した場合の増殖曲線を示した図である。
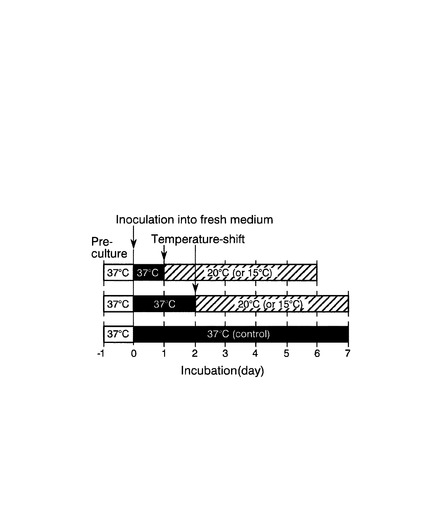
【図8】実施例3における各培養条件のスキーム図である。
【図9】実施例3において、各培養条件における培養液のOD600、培養液1mL当たりの乾燥菌体重量、および培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図10】実施例3において、脂肪酸組成の経時変化を示した図である。
【図11】実施例4における各培養条件のスキーム図である。
【図12】実施例4において、各培養条件における培養液のOD600、培養液1mL当たりの乾燥菌体重量、および培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図13】実施例5における各培養条件のスキーム図である。
【図14】実施例5において、各培養液の温度シフト時点および培養終了時点の培養液のOD600、培養終了時点の培養液1mL当たりの乾燥菌体重量、並びに培養終了時点の培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図15】実施例6における各培養条件のスキーム図である。
【図16】実施例6において、各培養液の培養終了時点の培養液のOD600、および培養終了時点の培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図17】実施例7において、各株のガスクロマトグラフ分析の結果のチャート図である。
【図18】実施例7において、各株の培養液1mL当たりの各種脂肪酸の、総脂肪酸含有量に対する含有割合(%)の結果を示した図である。
【図19】実施例8において、各形質転換体をスポットしたプレートをインキュベートした後の各プレートの写真図である。
【図20】実施例9において、各株の培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有比率の結果を示した図である。
【図21】参考例4において、各形質転換体を、20℃または30℃で培養した場合の増殖曲線を示した図である。
【図22】実施例10において、各株の脂肪酸組成の結果を示した図である。
【発明を実施するための形態】
【0010】
[FAH12遺伝子が導入された酵母の形質転換体(リシノール酸産生酵母)]
本発明におけるリシノール酸産生酵母は、FAH12遺伝子が導入された酵母の形質転換体である。酵母に導入するFAH12遺伝子は、オレイン酸の12位の炭素原子に水酸基を導入し得るΔ12−ヒドロキシラーゼ活性を有する酵素をコードする遺伝子であれば、特に限定されず、微生物由来であってもよく、植物由来であってもよい。具体的には、例えば、トウゴマのFAH12(RcFAH12)(GenBank ID.No.: AAC49010.1)をコードする遺伝子(RcFAH12遺伝子)、レスクェレラ・フェンドレリのFAH12(LfFAH12)(GenBank ID.No.: AAC32755.1)をコードする遺伝子(LfFAH12遺伝子)、麦角菌のFAH12(CpFAH12)(GenBank ID.No.: ACF37070.1)をコードする遺伝子(CpFAH12遺伝子)、フィサリア・リンドヘイメリ(Physaria lindheimeri)のoleate 12−hydroxylase(GenBank ID.No.:ABQ01458.1)をコードする遺伝子、アスペルギルス・ニガー(Aspergillus niger)のunnamed protein product(GenBank ID.No.:CAK37451.1)をコードする遺伝子、が挙げられる。なお、GenBankはNCBI(National Center for Biotechnology Information)のデータベースである。本発明では、RcFAH12遺伝子、LfFAH12遺伝子またはCpFAH12遺伝子が好ましく、Δ12−ヒドロキシラーゼ活性がより高いため、CpFAH12遺伝子がより好ましい。
【0011】
(酵母)
FAH12遺伝子が導入される宿主は、酵母であれば特に限定されず、出芽酵母(Saccharomyces cerevisiae、以下、S.セレビシエ)をはじめとするサッカロミセス(Saccharomyces)属酵母、シゾサッカロミセス(Schizosaccharomyces)属酵母、クリュイベロミセス(Kluyveromyces)属酵母、ピキア(Pichia)属酵母、カンジダ(Candida)属酵母が挙げられる。中でも、オレイン酸含有量が高いことから、シゾサッカロミセス属酵母であることが好ましい。シゾサッカロミセス属酵母としては、たとえば、S.ポンベ、シゾサッカロミセス・ジャポニカス(Schizosaccharomyces japonicus)、シゾサッカロミセス・オクトスポラス(Schizosaccharomyces octosporus)が挙げられ、S.ポンベが好ましい。
【0012】
また、宿主とする酵母は、野生型であってもよく、用途に応じて特定の遺伝子を欠失または失活させた変異型であってもよい。特定の遺伝子を欠失または失活させる方法としては、公知の方法を用いられる。具体的には、Latour法(Nucleic Acids Research誌、第34巻、e11ページ、2006年;国際公開第2007/063919号パンフレット等に記載)を用いることにより遺伝子を欠失させられる。また、変異剤を用いた突然変異分離法(酵母分子遺伝学実験法、1996年、学会出版センター)や、PCR(ポリメラーゼ連鎖反応)を利用したランダム変異法(PCR Methods Application誌、第2巻、28-33ページ、1992年)等により遺伝子の一部に変異を導入することにより該遺伝子を失活させられる。特定遺伝子を欠失または失活させたシゾサッカロミセス属酵母宿主としては、たとえば、国際公開第2002/101038号、国際公開第2007/015470号等に記載されている。
また、特定の遺伝子の削除または不活性化を行う部分はORF(オープンリーディングフレーム)部分であってもよく、発現調節配列部分であってもよい。特に好ましい方法は、構造遺伝子のORF部分をマーカー遺伝子に置換するPCR媒介相同組換え法(Yeast誌、第14巻、943-951ページ、1998年)による削除または不活性化の方法である。
【0013】
さらに宿主となる酵母には、形質転換体を選択するためのマーカーを有するものを用いることが好ましい。たとえば、ある遺伝子が欠落していることにより特定の栄養成分が生育に必須である宿主を使用することが好ましい。目的遺伝子配列を含むベクターにより形質転換をして形質転換体を作製する場合、ベクターにこの欠落している遺伝子(栄養要求性相補マーカー)を組み込んでおくことにより、形質転換体は宿主の栄養要求性が消失する。この宿主と形質転換体の栄養要求性の相違により、両者を区別して形質転換体を得られる。
たとえば、オロチジン5’−リン酸デカルボキシラーゼ遺伝子(ura4遺伝子)が欠失または失活してウラシル要求性となっている酵母を宿主とし、ura4遺伝子(栄養要求性相補マーカー)を有するベクターにより形質転換した後、ウラシル要求性が消失したものを選択することにより、ベクターが組み込まれた形質転換体を得られる。宿主において欠落により栄養要求性となる遺伝子は、形質転換体の選択に用いられるものであればura4遺伝子には限定されず、イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leu1遺伝子)等であってもよい。
【0014】
(発現カセット)
本発明におけるリシノール酸産生酵母は、FAH12遺伝子をコードする領域を含む構造遺伝子配列(FAH12遺伝子配列)、並びに該遺伝子を発現させるためのプロモーター配列およびターミネーター配列を含む発現カセットを、酵母に導入することにより作製できる。FAH12遺伝子配列としては、麦角菌等由来のFAH12をコードする遺伝子をそのまま用いてもよいが、宿主として用いる酵母内での発現量を増大させるために、前記遺伝子配列を、宿主での高発現遺伝子において使用頻度の高いコドンに改変することが好ましい。
【0015】
発現カセットとは、蛋白質を発現するために必要なDNAの組み合わせであり、該蛋白質をコードする構造遺伝子と宿主である酵母内で機能するプロモーターとターミネーターとを含む。
前記発現カセットには、さらに、5’−非翻訳領域、3’−非翻訳領域のいずれか1つ以上が含まれていてもよい。好ましい発現カセットは、FAH12遺伝子配列、プロモーター、ターミネーター、5’−非翻訳領域、3’−非翻訳領域を全て含む発現カセットである。さらに、栄養要求性相補マーカー等の遺伝子を有していてもよい。
【0016】
プロモーターとターミネーターは、宿主である酵母内で機能してFAH12を発現できるものであればよい。酵母内で機能するプロモーターとしては、酵母が本来有するプロモーター(転写開始活性が高いものが好ましい)や酵母が本来有しないプロモーター(ウイルス由来のプロモーターなど)を使用できる。プロモーターはベクター内に2種以上存在していてもよい。
シゾサッカロミセス属酵母が本来有するプロモーターとしては、たとえば、アルコールデヒドロゲナーゼ遺伝子プロモーター、チアミンの代謝に関与するnmt1遺伝子プロモーター、グルコースの代謝に関与するフルクトース−1、6−ビスホスファターゼ遺伝子プロモーター、カタボライト抑制に関与するインベルターゼ遺伝子のプロモーター(国際公開第99/23223号参照)、熱ショック蛋白質遺伝子プロモーター(国際公開第2007/26617号参照)などが挙げられる。
シゾサッカロミセス属酵母が本来有しないプロモーターとしては、たとえば、特開平5−15380号公報、特開平7−163373号公報、特開平10−234375号公報に記載されている動物細胞ウイルス由来のプロモーターが挙げられ、hCMVプロモーター、SV40プロモーターが好ましい。
シゾサッカロミセス属酵母内で機能するターミネーターとしては、シゾサッカロミセス属酵母が本来有するターミネーターやシゾサッカロミセス属酵母が本来有しないターミネーターを使用できる。ターミネーターはベクター内に2種以上存在していてもよい。
ターミネーターとしては、たとえば、特開平5−15380号公報、特開平7−163373号公報、特開平10−234375号公報に記載されているヒト由来のターミネーターが挙げられ、ヒトリポコルチンIのターミネーターが好ましい。
【0017】
(ベクター)
本発明におけるリシノール酸産生酵母は、具体的には、前記発現カセットを含むベクターにより宿主である酵母を形質転換することにより得られる。形質転換に用いるベクターは、環状DNA構造または線状DNA構造を有するベクターに、該発現カセットを組み込むことにより製造できる。該発現カセットが、宿主の細胞内で染色体外遺伝子として保持される形質転換体を作製する場合には、該ベクターは、宿主細胞内で複製されるための配列、即ち、自律複製配列(Autonomously Replicating Sequence: ARS)を含むプラスミドであることが好ましい。一方で、該発現カセットが、宿主細胞の染色体中に組み込まれた形質転換体を作製する場合には、該ベクターは、線状DNA構造であり、かつARSを有していないものとして、宿主細胞へ導入されることが好ましい。たとえば、該ベクターは、線状DNAからなるベクターであってもよく、宿主への導入時に、線状DNAに切り開くための制限酵素部位を備える環状DNA構造のベクターであってもよい。該ベクターがARSを有するプラスミドの場合、ARS部分を削除して線状DNA構造、またはARS部分を開裂させることによりARSの機能を失活させた線状DNA構造とした後、宿主へ導入できる。
【0018】
該ベクターは、形質転換体を選択するためのマーカーを有することが好ましい。該マーカーとしては、たとえば、ura4遺伝子(栄養要求性相補マーカー)、イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leu1遺伝子)が挙げられる。
【0019】
リシノール酸産生酵母の製造に用いられる前記発現カセットを含むベクターは、たとえば、外来構造遺伝子を導入するためのクローニングサイトを備える公知の発現ベクター中の該クローニングサイトに、FAH12遺伝子配列を挿入することによって製造できる。また、公知の発現ベクターに、FAH12遺伝子配列を含む発現カセットを組み込むことによっても製造できる。
【0020】
(形質転換体の製造方法)
本発明におけるリシノール酸産生酵母は、前記発現カセットを、染色体中に有するか、または、染色体外遺伝子として有する. 発現カセットを染色体中に有するとは、宿主細胞の染色体中の1カ所以上に発現カセットが組み込まれていることであり、染色体外遺伝子として有するとは、発現カセットを含むプラスミドを細胞内に有するということである。形質転換体の継代培養が容易であることから、該発現カセットを染色体中に有することが好ましい。
【0021】
形質転換方法は、公知の形質転換方法をいずれも用いられる。該形質転換方法としては、たとえば、酢酸リチウム法、エレクトロポレーション法、スフェロプラスト法、ガラスビーズ法など従来周知の方法や、特開2005−198612号公報記載の方法が挙げられる。また、市販の酵母形質転換用キットを用いてもよい。
【0022】
形質転換を行った後、通常は得られた形質転換体を選抜する。選抜方法としては、たとえば、以下に示す方法が挙げられる。前記栄養要求性マーカーにより形質転換体を選択できる培地によりスクリーニングし、得られたコロニーから複数を選択する。選抜した形質転換体に対してパルスフィールドゲル電気泳動法によるゲノム解析を行うことにより、染色体に組み込まれたベクターの数や発現カセットの数を調べられる。
【0023】
[リシノール酸産生酵母の培養方法]
酵母にFAH12を発現させた場合、通常の培養温度(30℃程度)では増殖阻害がかかるが、比較的高温で培養した場合には増殖阻害がかからない。高温条件下では、FAH12の発現が抑制されるか、または発現したFAH12のΔ12−ヒドロキシラーゼ活性が失活若しくは低下するためと推察される。本発明では、リシノール酸産生酵母をまず高温で培養した後、培養温度を低温にシフトさせることによって、該酵母内でリシノールを産生させる。はじめに高温で培養することにより、温度シフト後にもFAH12による増殖阻害の影響を抑制しつつ、FAH12によるリシノール酸産生を促進し得る。
すなわち、本発明のリシノール酸産生酵母の培養方法は、FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程とを有することを特徴とする。
【0024】
酵母の培養温度があまりに高い場合には、温度ストレスにより増殖がかえって低下する。リシノール酸産生酵母を35℃以上40℃未満で培養することにより、過度の高温ストレスを受けることなく、FAH12による増殖阻害作用を充分に抑制し得る。高温培養工程における培養温度は、35〜39℃が好ましく、35〜38℃がより好ましい。
【0025】
低温培養工程における培養温度を、高温培養工程よりも低い温度、すなわち35℃未満にすることで、増殖阻害作用が抑制されつつ、FAH12によるリシノール酸産生が促進される。但し、酵母の培養温度があまりに低い場合には、温度ストレスにより増殖がかえって低下する。そこで、低温培養工程における培養温度は、10〜33℃が好ましく、15〜33℃がより好ましく、20〜30℃がさらに好ましい。
【0026】
低温培養工程移行後にも、リシノール酸産生酵母が充分に増殖でき、培養液当たりのリシノール酸産生量を改善し得るため、酵母の増殖がある程度盛んな状態、特に対数増殖期にある段階で、高温培養工程から低温培養工程へ移行することが好ましい。このため、本発明においては、形質転換体の培養液のOD600が5.5以下の時点で、好ましくは5以下の時点で、より好ましくは4,4以下の時点で、高温培養工程から低温培養工程へ移行する。また、高温培養工程から低温培養工程へ移行する時点における形質転換体の培養液のOD600は、0.5以上であり、0.55以上であることがより好ましく、1.4以上であることがさらに好ましい。特に、リシノール酸産生酵母を、35℃以上40℃未満で、培養液のOD600が1.4〜4.4になるまで培養した後、当該培養液の温度を低温にシフトさせてさらに培養を続けることにより、より大量のリシノール酸を産生できる。培養液及びその希釈液のOD600は、例えば、分光光度計を使用して常法により測定できる。
【0027】
本発明のリシノール酸産生酵母の培養方法では、高温培養工程および低温培養工程のいずれでも、リシノール酸産生酵母は、培養温度以外は一般的な酵母の培養方法と同様にして培養できる。たとえば、培養液には、宿主とした酵母の培養に用いられる公知の培養培地を用いることができ、宿主が資化しうる炭素源、窒素源、無機塩類等を含有し、宿主の培養を効率良く行えるものであればよい。培養液としては、天然培地を用いてもよく、合成培地を用いてもよい。
【0028】
炭素源としては、たとえばグルコース、フルクトース、スクロース等の糖が挙げられる。
窒素源としては、たとえばアンモニア、塩化アンモニウム、酢酸アンモニウム等の無機酸または無機酸のアンモニウム塩、ペプトン、カザミノ酸が挙げられる。
無機塩類としては、たとえばリン酸マグネシウム、硫酸マグネシウム、塩化ナトリウムが挙げられる。
【0029】
培養には公知の細胞培養方法を用いることができ、たとえば振盪培養、攪拌培養等により行える。
培養は、回分培養であってもよく、連続培養であってもよい。また、培養時間は適宜決定できる。
【0030】
[リシノール酸の製造方法]
本発明のリシノール酸の製造方法は、本発明のリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とする。
培養されたリシノール酸産生酵母からのリシノール酸の抽出・精製は、溶剤抽出法等の一般的に脂肪酸の抽出・精製に使用される公知の手法により実施できる。例えば、リシノール酸産生酵母を培養した後、得られた培養液を遠心分離して菌体を回収し、該菌体をメタノール中でグラスビーズ等を用いて破砕しながら、クロロホルム/メタノール(1:2)を加えて抽出し、その後精製することにより、リシノール酸を精製できる。
また、リシノール酸は、ガスクロマトグラフィ法等の公知の手法で検出および定量できる。
【実施例】
【0031】
以下、実施例等を示して本発明を詳細に説明する。ただし、本発明は以下の記載によっては限定されない。
なお、以下の実施例等において、酵母の培養液および該希釈液のOD値は、分光光度計UV1600(島津製作所製)を用いて測定した。
【0032】
[酵母]
以下の実施例等において、S.ポンベは、ロイシンおよびウラシル要求株ARC010(遺伝子型:h− 、leu1−32、ura4−D18)(国際公開第2007/015470号パンフレット参照。)を用いた。また、S.セレビシエは、BY4741(遺伝子型:MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0)を用いた。
S.ポンベは、プラスミドを維持するために必要な選択圧に依存して、EMMSまたはEMMSドロップアウト培地中で培養した(Alfa et al., 1993 ,“A laboratory course manual.”, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.)。EMMSは通常の培養時に使用され、窒素が制限された最小培地(EMM−C/N3)がリシノール酸産生には使用された。発明者らによる従前の実験結果から、EMM−C/N3中で培養したほうが脂肪酸含有量が高くなることが示唆されていたためである。EMM培地は、0.5%(w/v)の塩化アンモニウム、2%(w/v)のグルコースを含有していた。一方、EMM−C/N3は、塩化アンモニウムの濃度は0.1%にまで低減されており、グルコース濃度が10%にまで増大していた。nmt1プロモーターの下、遺伝子発現を抑制するために、15μM(5μg/mL)のチアミンを使用した。
S.セレビシエは、プラスミドを維持するために必要な選択圧に依存して、最小合成培地(SD)、合成完全培地(SC)または合成完全ドロップアウト培地中で培養した(Sherman, et.al., 1986, “Methods in yeast genetics. ”, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.)。最小合成培地またはSCは、0.17%(w/v)のアミノ酸およびアンモニウム塩不含有イーストニトロジェンベース(Bactoyeast nitrogen base、Difco社製)、2%(w/v)のグルコース、および0.5%(w/v)の硫酸アンモニウムを含有していた。窒素が制限された最小培地(NSD)では、硫酸アンモニウム含有量が0.1%にまで低減されており、グルコース濃度が10%にまで増大していた(Yazawa, et.al., 2007,“Heterologous production of dihomo-gamma-linolenic acid in yeast Saccharomyces cerevisiae.”, Appl Env Microbiol, vol.73, pp.6965-6971)。
【0033】
[プラスミド構築]
RcFAH12遺伝子、LfFAH12遺伝子、およびCpFAH12遺伝子のコード領域を含む酵母の形質転換用プラスミドを構築した。DNA操作の標準的な技術は、SambrookおよびRussellの方法(“Molecular cloning: A laboratory manual. 3rd ed.”, 2001, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.)に準じて行った。また、プラスミド構築は、大腸菌DH5α(F−endA1、hsdR17、supE44、thi−1、recA1、gyrA96、relA1、Δ(argF−lacZ)U169、Φ80、Δ(lacZ)M15)株を用いて行った。当該DH5α株は、アンピシリンまたはカナマイシンを添加したLB培地(Luria-Bertani broth)中で培養した。
【0034】
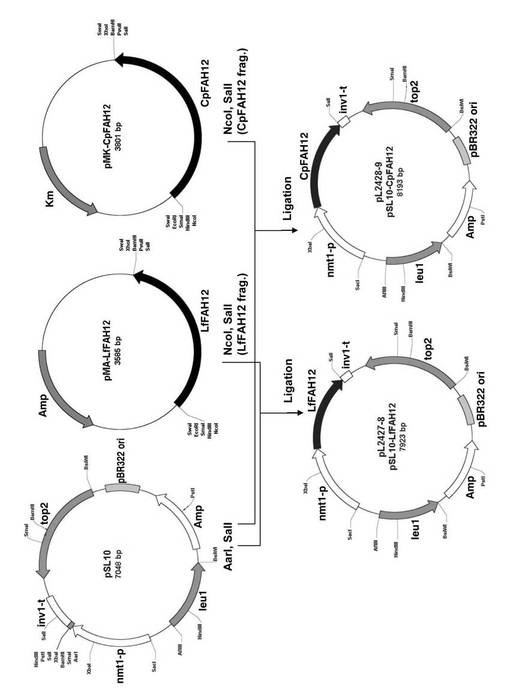
CpFAH12遺伝子のDNA配列(Meesapyodsuk and Qiu, 2008, Plant Physiology, vol.147, pp.1325−1333)は、S.ポンベにおいて好ましい使用頻度のコドンに改変した後、GENEART社(Regensburg, Germany)に依頼して化学合成した。合成されたCpFAH12のORFは、5’末端と3’末端にそれぞれ人工的に付加されていた制限酵素部位を使用して、大腸菌のプラスミドpMK−CpFAH12から、制限酵素NcoIおよびSalIで二重消化したフラグメントとして切り出し、pL2428−9(略してpSL10−CpFAH12)を構築するために、マルチクローニングサイトの制限酵素AarIおよびSalIで二重消化したプラスミドpSL10に組み込んだ。合成されたプラスミドをpSL10−CpFAH12とした。pSL10は、leu1マーカーを有する、染色体へ組み込まれるタイプのプラスミドであり、pSL6プラスミド(Alimjan et al., Appl Microbiol Biotechnol, 2010, vol.85, pp.667−677)のhCMVプロモーター(hCMV−p)をnmt1プロモーターに置換し、LPIターミネーター(LPI−ter.)をinv1ターミネーターに置換したプラスミドである。pSL10の塩基配列を配列番号1に示す。S.ポンベにpSL10−CpFAH12を導入した形質転換体中では、チアミン非存在時には、nmt1プロモーターの制御下でCpFAH12が発現したが、チアミン存在時には発現は抑制された。
pSL10−CpFAH12(pL2428−9)と同様にして、LfFAH12遺伝子のORFを組み込んだpL2427−8 (略してpSL10−LfFAH12)を構築した。
図1に、pSL10−CpFAH12およびpSL10−LfFAH12のベクターの構築手順を模式的に示した。なお、同様な手法によりpSL10−RcFAH12も作製できる。
【0035】
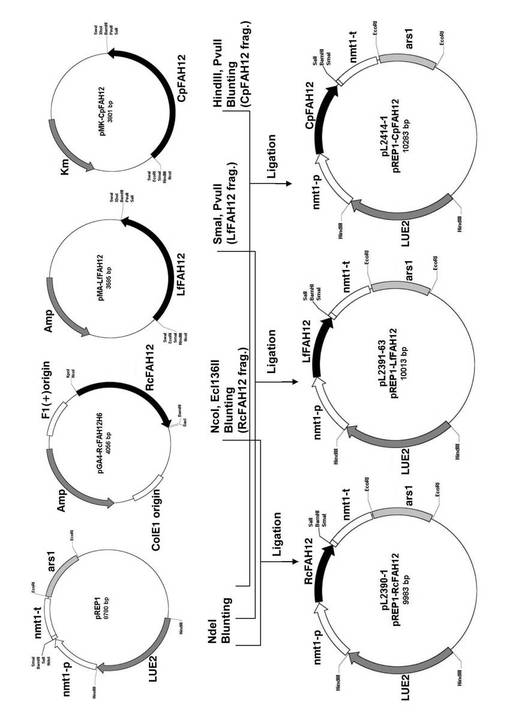
また、pMK−CpFAH12から、制限酵素HindIIIおよびPvuIIで二重消化したフラグメントとしてCpFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。合成されたプラスミドをpL2414−1(略してpREP1−CpFAH12)とした。pREP1は、S.セレビシエのLEU2マーカーを有する染色体外に存在するタイプのプラスミドである。S.ポンベにpREP1−CpFAH12を導入した形質転換体中では、チアミン非存在時には、nmt1プロモーターの制御下でCpFAH12が発現したが、チアミン存在時には発現は抑制された。
pL2390−1(略してpREP1−RcFAH12)は、pGA4−RcFAH12H6を制限酵素NcoIおよびEcl136IIで二重消化したフラグメントとしてRcFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。pL2391−63(略してpREP1−LfFAH12)は、pMA−LfFAH12を制限酵素SmaIおよびPvuIIで二重消化したフラグメントとしてLfFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。
図2に、pREP1−RcFAH12、pREP1−LfFAH12、およびpREP1−CpFAH12のベクターの構築手順を模式的に示した。
【0036】
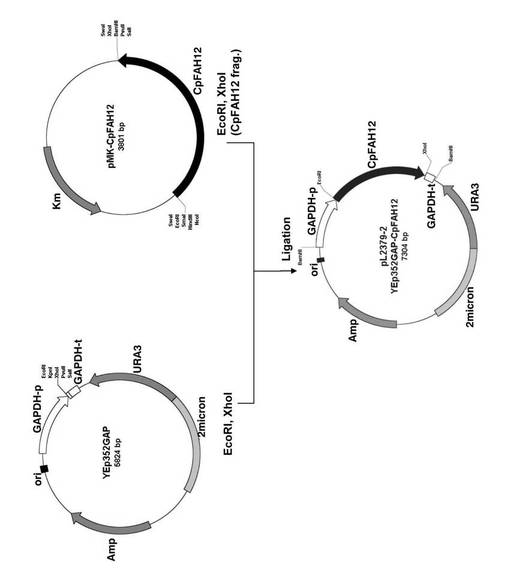
また、pMK−CpFAH12から、制限酵素EcoRIおよびXhoIで二重消化したフラグメントとしてCpFAH12を切り出し、pL2379−2を構築するために、マルチクローニングサイトの制限酵素EcoRIおよびXhoIで二重消化したプラスミドYEp352GAPに組み込んだ。S.セレビシエにpL2379−2(略してYEp352−CpFAH12)を導入した形質転換体中では、解糖系のGAPDHプロモーターの制御下でCpFAH12が発現した。
図3に、YEp352−CpFAH12のベクターの構築手順を模式的に示した。なお、同様な手法によりYEp352−RcFAH12も作製できる。
【0037】
[形質転換体の作製]
酵母細胞は、Frozen−EZ Yeast Transformation II kit (Zymo Research, CA, USA)を用いて、製造者により推奨されるプロトコールに従って形質転換した。
たとえば、CpFAH12遺伝子を染色体中に組み込む形で導入したS.ポンベの形質転換体の作製は、以下のようにして行った。すなわち、酵母の第2染色体のleu1座にpSL10−CpFAH12を組み込むため、pSL10−CpFAH12は制限酵素BsiWIによって消化し、得られた線状プラスミドをS.ポンベARC010−1株に導入し、leu1+の形質転換体を選抜した。
一方で、CpFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体は、pREP1−CpFAH12をそのままS.ポンベARC010−1株に導入し、LEU2+の形質転換体(すなわち、ロイシン要求性のない形質転換体)を選抜することによって作製した。
RcFAH12遺伝子またはLfFAH12遺伝子を染色体中または染色体外に導入したS.ポンベの形質転換体は、CpFAH12遺伝子を染色体中または染色体外に導入したS.ポンベの形質転換体と同様にして作製した。
【0038】
[参考例1]
S.ポンベおよびS.セレビシエの脂肪酸含有量を測定した。
具体的には、S.ポンベの場合は、EMM−C/N3(ロイシンおよび/またはウラシル無含有培地)中で、30℃で一晩前培養し、該前培養物を新しいEMM−C/N3に接種し、40rpmで振とう培養した。S.セレビシエの場合は、NSD培地を用いた。
得られた培養物の総脂肪酸含有量は、ガスクロマトグラフ分析によるKainouらの方法(2006, Yeast, vol.23, pp.605-612)に準じて行った。具体的には、以下に示す通りである。まず、脂肪酸分析は、温度制御環境下(160〜234℃、昇温速度:4.5℃/min)で、TC−70キャピラリーカラム(30m×0.25mm(内径)、GL Sciences製)を備えたガスクロマトグラフ(GC2010、島津製作所製)に0.2μLの培養液を投与して実施した。脂肪酸組成は、各ピークの面積に基づいて算出され、含有量は、標準としたヘプタデカン酸メチル(C17:0)と比較することによって決定した。
【0039】
測定結果を図4に示す。縦軸は全脂肪酸に対する各脂肪酸の含有量割合(%)である。S.セレビシエでは、オレイン酸(C18:1)は30%強含まれていたが、パルミトレイン酸(C16:1)が約45%と最も多かった。一方、S.ポンベでは、80%強がオレイン酸(C18:1)であり、その他の脂肪酸含有量はいずれも非常に少なかった。
【0040】
[参考例2]
pREP1、pREP1−CpFAH12、pREP1−RcFAH12、またはpREP1−LfFAH12を導入したS.ポンベの形質転換体を、それぞれチアミン存在下および非存在下で培養し、増殖能を調べた。
具体的には、まず、チアミン含有EMM−C/N3−LEU(EMM−C/N3からロイシンが除去された培地)中、30℃で一晩前培養した前培養液を、チアミン不添加のEMM−C/N3−LEUのプレート(アガー平板培地)またはチアミン含有EMM−C/N3−LEUのプレートに塗布し、30℃で4日間培養し、コロニーの形成の有無や、形成されたコロニーのサイズを調べた。
空ベクターであるpREP1を導入した形質転換体をチアミン含有プレートに塗布した場合に形成されたコロニーのサイズを基準として、その他の形質転換体の増殖能を評価した。評価結果を表1に示す。表1中、「+++」は生育が正常であり、「++」は生育が若干悪く、「+」は生育が悪く、「−」は増殖できなかったことを意味する。また、「遺伝子発現OFF(チアミン有り)」はチアミン含有EMM−C/N3−LEUプレートの結果を、「遺伝子発現ON(チアミン無し)」はチアミン不添加のEMM−C/N3−LEUプレートの結果を、それぞれ示す。
【0041】
【表1】

【0042】
RcFAH12を発現させた場合には、外来蛋白質を発現させていない場合と同程度の増殖能であったが、LfFAH12を発現させた形質転換体は、RcFAH12を発現させた形質転換体よりも増殖能が低下しており、CpFAH12を発現させた形質転換体は非常に増殖能が低下していることがわかった。つまり、LfFAH12やCpFAH12を発現させることによって、増殖能が低下した。
【0043】
[参考例3]
pREP1−LfFAH12を導入したS.ポンベの形質転換体を培養し、リシノール酸をはじめとする脂肪酸の含有量を調べた。
具体的には、まず、チアミン含有EMM−C/N3−LEU中、30℃で一晩前培養した前培養液を、EMM−C/N3−LEU中に接種し、30℃、40rpmで5日間振とう培養した。得られた培養物の各脂肪酸の含有量を、参考例1と同様にしてガスクロマトグラフ分析によって測定した。測定の結果得られたチャート図を図5に示す。該形質転換体では、リシノール酸の標準サンプルと同じ位置にピークが検出された。すなわち、該形質転換体ではリシノール酸が生産されていることが確認できた。
【0044】
[実施例1]
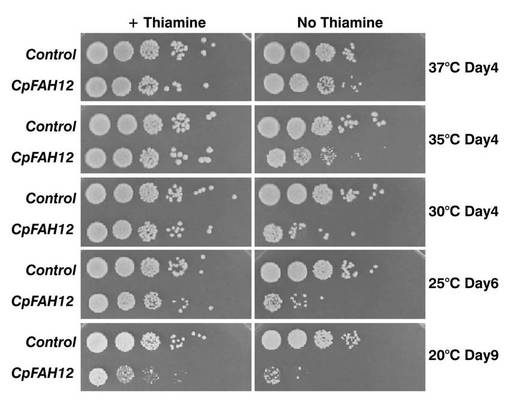
CpFAH12遺伝子を染色体中に組み込んだS.ポンベの形質転換体(以下、CpFAH12インテグラント株)の各培養温度における増殖能を調べた。
具体的には、CpFAH12インテグラント株にura4マーカーを含む空ベクターpREP2を導入した形質転換体(CpFAH12株)と、S.ポンベにpSL10およびpREP2を導入した形質転換体(コントロール株)を、それぞれチアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一晩、培養液のOD600が約4になるまで前培養した。該前培養液を同種の新しい培地で希釈し、さらに、10倍希釈系列を調整し、各10μLずつを、15μMのチアミン含有EMM−C/N3−URA,LEUプレート(発現抑制条件)またはチアミン不添加のEMM−C/N3−URA,LEUプレート(発現誘導条件)上にスポットした。該プレートを37℃、35℃、30℃、25℃、または20℃で4〜9日間インキュベートした。
インキュベート後の各プレート表面の写真図を図6に示す。図中、「Control」はコントロール株の結果であり、「CpFAH12」はCpFAH12株の結果を示す。また、図中、白い部分がコロニーである。チアミン含有プレート(図中、「+Thiamine」)では、20℃、9日間培養した場合のみ、CpFAH12株がコントロール株よりもやや増殖能が低かったものの、その他の培養条件では両形質転換体の増殖能に特段の差はなかった。一方、参考例2ではCpFAH12を発現させた株ではほとんど増殖が観察されなかったにも関わらず、チアミン不含有プレート(図中、「No Thiamine」)では、濃い濃度でスポットとした場合に、全ての培養条件でCpFAH12株の増殖が確認できた。但し、37℃、4日間培養した場合を除き、CpFAH12株のほうがコントロール株よりも明らかに増殖能が低かった。すなわち、CpFAH12の発現誘導条件下のCpFAH12インテグラントの増殖能は培養温度に影響され、37℃で前培養した後に培養温度を低下させた場合には、増殖能は低下するものの、増殖自体は可能であること、および37℃ではほとんど低下しないことがわかった。
【0045】
[実施例2]
実施例1で用いたCpFAH12株およびコントロール株の、30℃または37℃の液体培地中の増殖能を調べた。
具体的には、CpFAH12株およびコントロール株を、チアミン不添加のEMM−C/N3−URA,LEU中、37℃で一晩前培養した。該前培養物を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、30℃または37℃で36時間、攪拌培養した。630nmの濁度を自動検出器(Bio−Plotter、東洋測器製)でモニターすることにより、液体培地中の増殖曲線を描いた。図7に増殖曲線を示す。図7(A)は30℃で培養した結果であり、図7(B)は37℃で培養した結果である。図中、三角がCpFAH12株の増殖曲線を、円がコントロール株の増殖曲線を、それぞれ示す。37℃で培養した場合よりも、30℃で培養した場合には、CpFAH12株はコントロール株よりも明らかに増殖が抑制されていた。
【0046】
[実施例3]
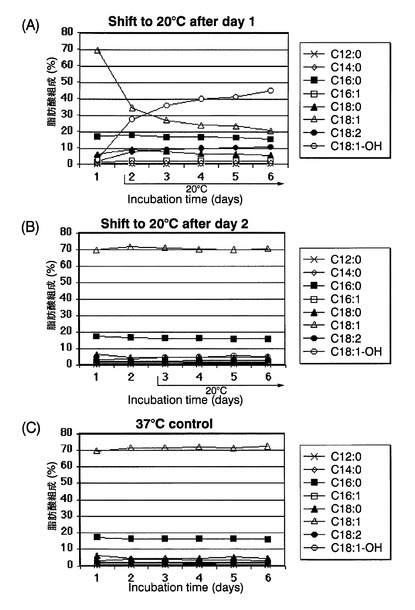
37℃から20℃または15℃に培養温度をシフトさせるタイミングの、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
図8は、評価方法のスキーム図である。まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、最初に37℃で1日間(OD600が5、対数増殖期後期)または2日間(OD600が6、定常増殖期初期)培養し、その後20℃または15℃に培養温度をシフトさせて5日間培養した。対照として、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、温度シフトせずに37℃で7日間培養した(図8の下段)。液体培養は全て、40rpmで振とう培養した。
【0047】
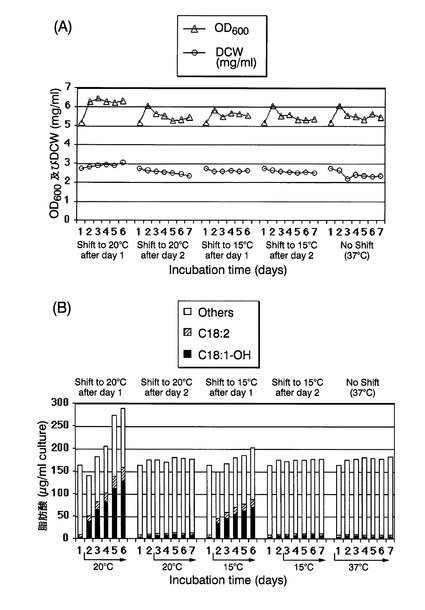
本培養開始後1日経過ごとに、培養液の一部をサンプリングし、培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液のOD600は実施例2と同様にして、各種脂肪酸の含有量は参考例1と同様にして、それぞれ測定した。培養液のOD600および培養液1mL当たりの乾燥菌体重量の測定結果を図9(A)に、各種脂肪酸の含有量の測定結果を図9(B)に、それぞれ示す。図9(B)中、「C18:1−OH」はリシノール酸を、「C18:2」はリノレン酸を、「Others」はその他の脂肪酸の総和を、それぞれ示す。培養液のOD600および培養液1mL当たりの乾燥菌体重量は、培養温度の温度シフトによってはあまり差がなかった。しかし、各種脂肪酸の含有量は、温度シフトせずに37℃で培養し続けた場合、37℃で2日間培養した後に20℃または15℃にシフトした場合には、リシノール酸の含有量は非常に低かった。これに対して、37℃で1日間培養した後に20℃または15℃にシフトした場合には、培養時間の経過とともにリシノール酸の含有量が顕著に増大していた。該結果から、培養温度が37℃の場合、および増殖が定常状態になった後に低温へ温度シフトした場合には、リシノール酸が産生されないことが分かった。培養温度が37℃の場合等でリシノール酸産生が行われないのは、CpFAH12が発現しないため、またはCpFAH12の活性が非常に低下するためと推察される。
【0048】
さらに、図10に、脂肪酸組成の経時的変化を示す。図10(A)は37℃で1日間培養した後に20℃にシフトして5日間培養した場合、図10(B)は37℃で2日間培養した後に20℃にシフトして5日間培養した場合、図10(C)は37℃で7日間培養した場合の結果である。37℃で2日間培養した後に20℃にシフトした場合には、37℃で7日間培養した場合と同様、脂肪酸組成はほとんど経時的変化がなかった。これに対して37℃で1日間培養した後に20℃にシフトした場合には、培養時間の経過と共にオレイン酸含有割合が低下し、逆にリシノール酸含有割合が増大した。その他の脂肪酸の含有割合は、ほとんど経時的変化がなかった。
【0049】
[実施例4]
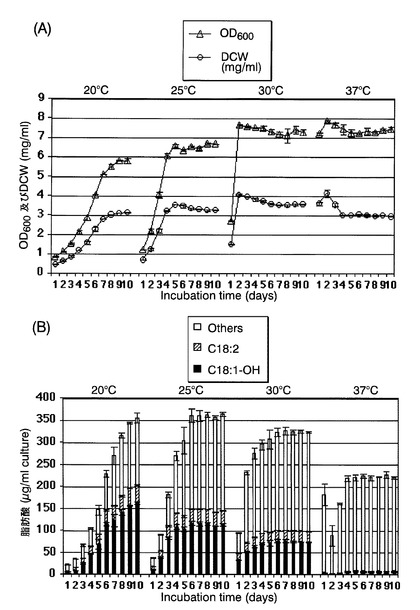
37℃から20℃、25℃または30℃への培養温度シフトの、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図11に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、20℃、25℃または30℃で10日間、40rpmで振とう培養した(図11の上段)。対照として、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、温度シフトせずに37℃で10日間、40rpmで振とう培養した(図11の下段)。
実施例3と同様に、本培養開始後1日経過ごとに、培養液の一部をサンプリングし、培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液のOD600および培養液1mL当たりの乾燥菌体重量の測定結果を図12(A)に、各種脂肪酸の含有量の測定結果を図12(B)に、それぞれ示す。図12(B)中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。
【0050】
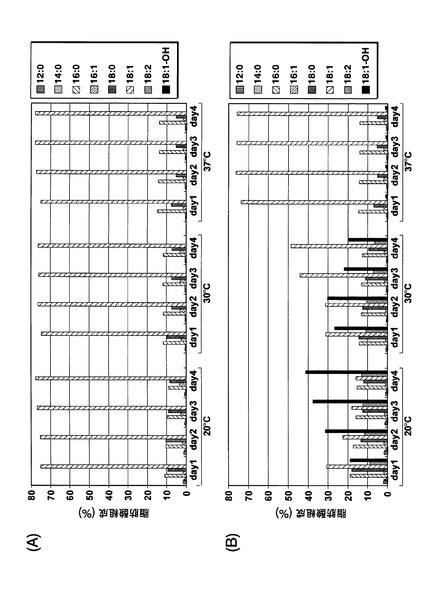
最終的なリシノール酸産生量は、本培養を20℃で実施した場合が、25℃または30℃で実施した場合よりも多くなることが確認された。しかし、本培養を25℃で行うほうが、利点があると考えられる。細胞増殖は25℃のほうが良好であるため、本培養開始4日目では、25℃の培養条件のほうが20℃よりもリシノール酸の産生量が高かったためである。具体的には、培養液当たりのリシノール酸産生量は、20℃で10日間培養後には163.2μg/mLであったが、20℃で4日間培養後には48.2μg/mLでしかなかった。これに対して25℃で4日間培養後には108.8μg/mLであり、該生産量は、20℃で10日間培養した場合の66%であった。さらにコストの点から、冷却コストを低減でき、かつ培養日数を10日間から4日間に短縮することで培養コストを低減できるため、細胞増殖は20℃よりも25℃で行うほうが好ましい。このように、冷却コストを低減し、培養時間を短縮することで製造コストを低減できるため、工業上の観点からは、25℃培養のほうが20℃培養よりも好ましい。
【0051】
[実施例5]
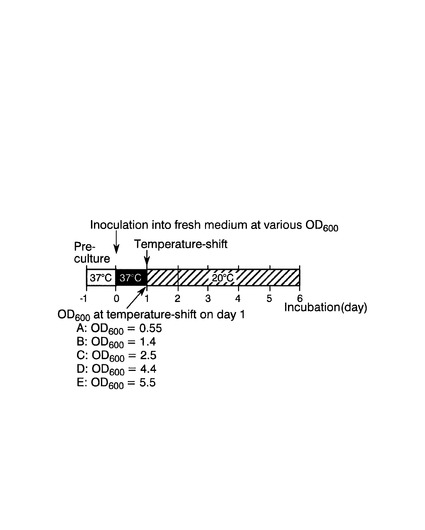
37℃から20℃に培養温度をシフトさせる時点の酵母の増殖状態の、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図13に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.01〜0.16の間様々な値になるように希釈した5つの本培養液を調製後、37℃で1日間、40rpmで振とう培養した後、培養液の温度を20℃にシフトし、シフト後さらに5日間、40rpmで振とう培養した(図13の上段)。5つの培養液の温度シフト時のOD600は、それぞれ、0.55(培養液A)、1.4(培養液B)、2.5(培養液C)、4.4(培養液D)および5.5(培養液E)であった。対照として、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、温度シフトせずに37℃で6日間、40rpmで振とう培養した(図13の下段)。
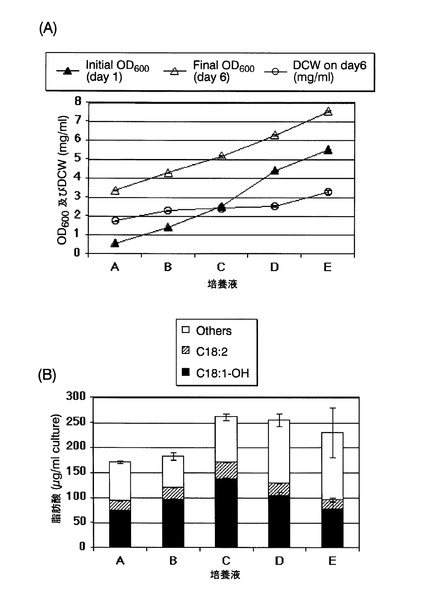
実施例3と同様にして、本培養終了時点の培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。温度シフト開始時点および培養終了時点の培養液のOD600、並びに培養終了時点の培養液1mL当たりの乾燥菌体重量の測定結果を図14(A)に、培養終了時点の培養液1mL当たりの各種脂肪酸の含有量の測定結果を図14(B)に、それぞれ示す。図14(B)中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。さらに、総脂肪酸含有量に対する各脂肪酸の含有量比(質量%)を表2に示す。図14および表2の各測定値は、独立した2回の試行を行い、その平均値である。
【0052】
【表2】
【0053】
図14(B)および表2に示す通り、培養液A〜Eの全てでリシノール酸の産生が確認されたが、温度シフト時点の培養液のOD600が1.4〜4.4である培養液B〜Dが、培養液AおよびEよりもリシノール酸産生量が多かった。該結果から、酵母が対数増殖期にある時点で温度シフトを行うことにより、より大量のリシノール酸を産生させ得ることが示唆された。
【0054】
[実施例6]
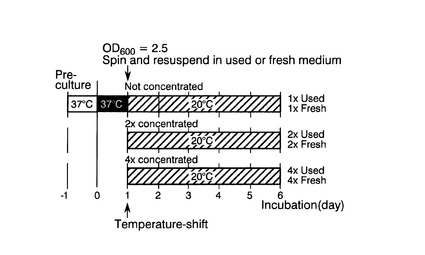
37℃から20℃に培養温度をシフトさせる時点の培養液の濃度の、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図15に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.07になるように希釈した後、37℃で培養液のOD600が2.5になるまで40rpmで振とう培養した後、培養液を遠心分離処理して菌体を回収した後、遠心分離処理前の菌体濃度の1倍、2倍、または4倍となるように、元々の培地または新しい同種の培地に懸濁して培養液を調製した。該培養液の温度を20℃にシフトし、シフト後さらに5日間、40rpmで振とう培養した。
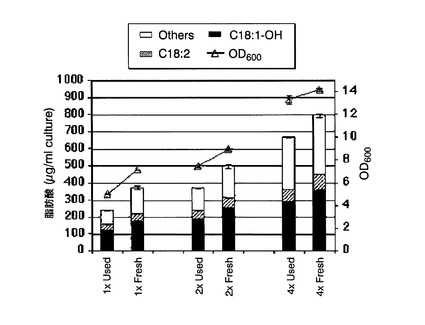
実施例3と同様にして、本培養終了時点の培養液のOD600、および培養液1mL当たりの各種脂肪酸の含有量を測定した。測定結果を図16に示す。図16中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。また、「1×Used」、「2×Used」および「4×Used」は、元々の培地で菌体濃度が遠心分離処理前の菌体濃度の1倍、2倍、または4倍になるように調製した培養液の結果であり、「1×Fresh」、「2×Fresh」および「4×Fresh」は、新しい培地で菌体濃度が遠心分離処理前の菌体濃度の1倍、2倍、または4倍になるように調製した培養液の結果である。
【0055】
遠心分離処理後の培養液の調製に新しい培地を用いたほうが、元々の培地を用いるよりも細胞増殖速度が速く、よって培養液1mL当たりのリシノール酸産生量も多くなった。また、遠心分離処理後の菌体の濃縮倍率が大きいほど、培養液1mL当たりのリシノール酸産生量も多くなった。しかし、総脂肪酸全体に占めるリシノール酸の含有割合は、各培養液において特に大きな差はなかった。
【0056】
[実施例7]
37℃から20℃または30℃への培養温度シフトの、実施例1で用いたCpFAH12株およびコントロール株のリシノール酸産生に対する影響を評価した。
まず、CpFAH12株およびコントロール株をそれぞれ、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、20℃、30℃または37℃に培養温度をシフトさせて4日間培養した。本培養開始後1日経過ごとに、培養液の一部をサンプリングし、参考例1と同様にして、培養液1mL当たりの各種脂肪酸の含有量を測定した。
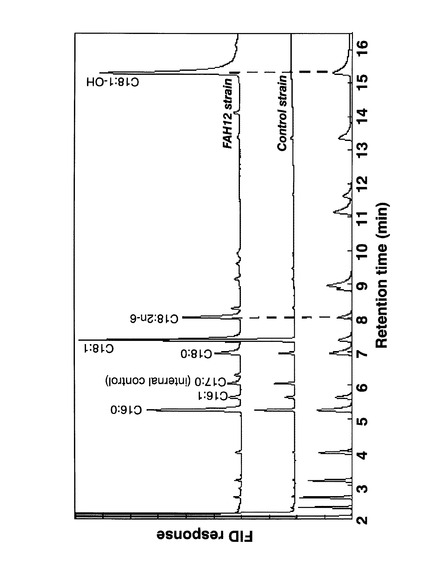
本培養を20℃で行った場合の各株のガスクロマトグラフ分析の結果のチャート図を図17に示す。CpFAH12株では、リシノール酸の標準サンプルと同じ位置にピークが検出された。該ピークは、CpFAH12を発現していないコントロール株では観察されなかった。
培養液1mL当たりの各種脂肪酸の、総脂肪酸含有量に対する含有割合(%)の結果を図18に示す。図18(A)はコントロール株の結果であり、図18(B)はCpFAH12株の結果である。CpFAH12を発現していないコントロール株では、本培養の温度に関わらず、脂肪酸組成は全てほぼ同様であった。これに対してCpFAH12株では、37℃で培養した場合にはコントロール株の結果とほぼ変わらなかったのに対して、20℃および30℃で培養した場合には、オレイン酸の含有割合の低下に伴い、リシノール酸の含有割合が増大していた。20℃の場合には、培養日数が長くなるにつれてリシノール酸含有割合が増大していた。30℃の場合には、培養日数が2日目でリシノール酸の含有割合はピークを迎えていた。
【0057】
[実施例8]
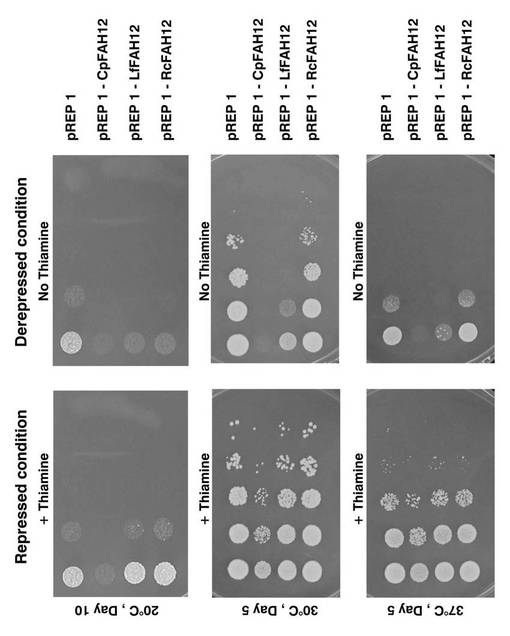
pREP1、pREP1−CpFAH12、pREP1−RcFAH12、またはpREP1−LfFAH12を導入したS.ポンベの形質転換体を、それぞれチアミン存在下および非存在下で培養し、各培養温度における増殖能を調べた。
まず、各形質転換体をそれぞれ、チアミンが添加されていないEMM−C/N3−LEU中、37℃で一日間前培養した。次いで、該前培養液を同種の新しい培地で希釈し、さらに、10倍希釈系列を調整し、各10μLずつを、15μMのチアミン含有EMM−C/N3−LEUプレート(発現抑制条件)またはチアミン不添加のEMM−C/N3−LEUプレート(発現誘導条件)上にスポットした。該プレートを20℃で10日間、30℃で5日間、または37℃で5日間インキュベートした。インキュベート後の各プレート表面の写真図を図19に示す。図中、白い部分がコロニーである。図中、「+Thiamine」がチアミン含有プレートであり、「No Thiamine」がチアミン不含有プレートである。
チアミン不含有プレートでは、培養温度にかかわらず、pREP1−CpFAH12を導入した形質転換体およびpREP1−LfFAH12を導入した形質転換体では、pREP1を導入した形質転換体よりも増殖能が低い傾向が観察された。
【0058】
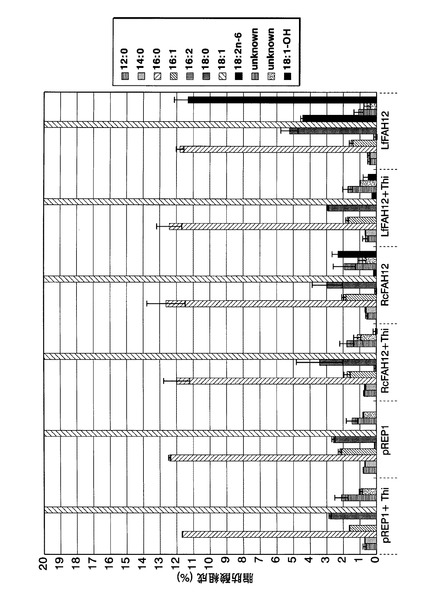
[実施例9]
RcFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体(以下、pREP1−RcFAH12導入株)およびLfFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体(以下、pREP1−LfFAH12導入株)の30℃で培養した場合の脂肪酸組成を調べた。対照として、pREP1を導入したS.ポンベの形質転換体(以下、pREP1導入株)を30℃で培養した場合の脂肪酸組成を調べた。
まず、各株をチアミンが添加されていないEMM−C/N3−LEU中、30℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地またはチアミン含有EMM−C/N3−LEUで、OD600が0.05になるように希釈した後、30℃で7日間、40rpmで振とう培養した。参考例1と同様にして、培養終了後の培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有比率の結果を図20に示す。図20中、「pREP1」、「RcFAH12」、および「LfFAH12」はそれぞれ、pREP1導入株、pREP1−RcFAH12導入株、およびpREP1−LfFAH12導入株をチアミンが添加されていないEMM−C/N3−LEU中で本培養した結果を示し、「pREP1+Thi」、「RcFAH12+Thi」、および「LfFAH12+Thi」はそれぞれ、pREP1導入株、pREP1−RcFAH12導入株、およびpREP1−LfFAH12導入株をチアミン含有EMM−C/N3−LEU中で本培養した結果を示す。
【0059】
図20に示すように、pREP1−RcFAH12導入株およびpREP1−LfFAH12導入株では、チアミン含有培地ではpREP1導入株と同様、リシノール酸はほとんど含有されていなかったが、チアミンが添加されていない培地では、オレイン酸含有割合が低下し、リシノール酸の含有割合が上昇していた。すなわち、CpFAH12遺伝子を導入した形質転換体と同様、RcFAH12遺伝子またはLfFAH12遺伝子を導入した形質転換体では、30℃で培養した場合に、リシノール酸が産生されることがわかった。
【0060】
[参考例4]
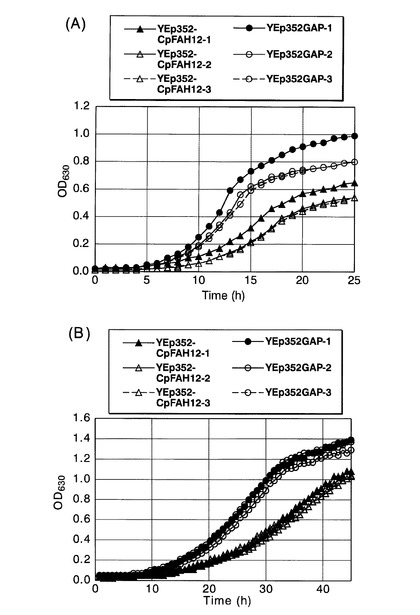
CpFAH12遺伝子を染色体外遺伝子として導入したS.セレビシエの形質転換体の増殖能を調べた。
具体的には、該S.セレビシエにYEp352−CpFAH12を導入した形質転換体(CpFAH12株)と、S.セレビシエに空ベクターYEp352GAPを導入した形質転換体(コントロール株)を、それぞれSC−URA中、30℃で一晩前培養した。該前培養物を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、20℃または30℃で攪拌培養した。630nmの濁度を自動検出器(Bio−Plotter、東洋測器製)でモニターすることにより、液体培地中の増殖曲線を描いた。それぞれ独立した試行を3回行った。図21に増殖曲線を示す。図21(A)は30℃で培養した結果であり、図21(B)は20℃で培養した結果である。図中、三角がCpFAH12株の増殖曲線[YEp352−CpFAH12−1、YEp352−CpFAH12−2、YEp352−CpFAH12−3]を、円がコントロール株の増殖曲線[YEp352GAP−1、YEp352GAP−2、YEp352GAP−3]を、それぞれ示す。いずれの培養温度においても、コントロール株よりもCpFAH12株のほうが、明らかに増殖が抑制されていた。すなわち、CpFAH12株の増殖のダブリングタイムは、コントロール株よりも1.4倍長くなっていた。
CpFAH12遺伝子を染色体中に組み込んだS.セレビシエの形質転換体は、CpFAH12遺伝子を染色体中に組み込んだS.ポンベの形質転換体と同様、CpFAH12を発現させることによって増殖が抑制されていた。つまり、CpFAH12遺伝子をS.セレビシエに導入することによって、S.ポンベに導入した場合と同様、リシノール酸を産生できることが示唆された。
【0061】
[実施例10]
CpFAH12遺伝子を染色体外遺伝子として導入したS.セレビシエの形質転換体を培養し、リシノール酸の産生能を調べた。対照として、S.セレビシエにYEp352GAPを導入した形質転換体を用いた。
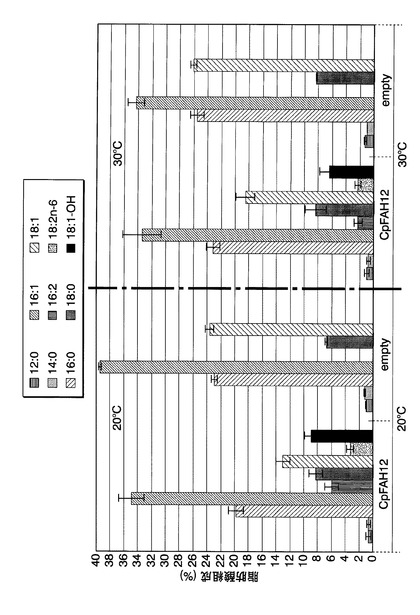
まず、各株をNSD−URA中、20℃または30℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、20℃または30℃で5日間、40rpmで振とう培養した。参考例1と同様にして、培養終了後の培養液1mL当たりの各種脂肪酸の含有量を測定した。脂肪酸組成(培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有割合)の結果を図22に示す。図22中、「CpFAH12」および「empty」はそれぞれ、CpFAH12を染色体外遺伝子として導入した形質転換体、およびS.セレビシエにYEp352GAPを染色体外遺伝子として導入した形質転換体を本培養した結果を示す。
図22に示すように、CpFAH12遺伝子を導入したS.セレビシエを20℃または30℃で培養した場合に、リシノール酸が産生されることがわかった。
【産業上の利用可能性】
【0062】
本発明のリシノール酸産生酵母の培養方法およびリシノール酸の製造方法により、リシノール酸を、酵母の形質転換体を用いて効率よく生産できるため、特に、リシノール酸またはひまし油が原料として使用されている化成品、塗料・印刷インキ、化粧品、医薬品、潤滑油等の製造分野において好適に用いられる。
【技術分野】
【0001】
本発明は、酵母によりリシノール酸を産生させる方法に関する。より詳細には、FAH12遺伝子が導入された酵母の形質転換体を培養することにより、該形質転換体にリシノール酸を産生させる方法に関する。
【背景技術】
【0002】
現在、ウレタン製品等の化成品の原料は、専ら石油資源に頼っている。地球温暖化対策により、石油資源に代わる代替材料が模索されており、リシノール酸が注目されている。リシノール酸は、トウゴマ(Ricinus communis)を絞ったひまし油の80〜90%を占めており、ひまし油から精製したものが使用されている。しかし、トウゴマにはリシンという猛毒の蛋白質が含まれており、取り扱いに注意を要する上、植物資源であるため、安定供給には不安がある。そこで、ひまし油以外からリシノール酸を安全かつ安定して製造する方法の開発が望まれている。
【0003】
リシノール酸は、鎖長が炭素数18であり、不飽和結合が1つと12位に水酸基を持つ脂肪酸であり、Δ12−ヒドロキシラーゼにより、オレイン酸の12位に水酸基を導入することによって生産できる。野生型の酵母はΔ12−ヒドロキシラーゼを有していないため、リシノール酸を産生することはできない。そこで、本発明者らは、分裂酵母シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe、以下、S.ポンベ)が、脂肪酸組成としてオレイン酸の含有量が8割程度と高いことに着目して、S.ポンベに異種生物由来のΔ12−ヒドロキシラーゼ遺伝子を導入し、リシノール酸の生産を試みた。この結果、S.ポンベの形質転換体においてリシノール酸が生産されることが確認されたが、リシノール酸の生産量が多くなるほど宿主の増殖が遅くなることも分かった(非特許文献1参照。)。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】石川大輔、他8名、日本分子生物学会年会講演要旨集、2009年、第32巻、第2号、第70ページ。
【発明の概要】
【発明が解決しようとする課題】
【0005】
そこで本発明の目的は、リシノール酸産生酵母を、増殖性とリシノール産生性のバランスをとりながら効率よく培養する方法、およびリシノール酸の製造方法を提供することにある。
【課題を解決するための手段】
【0006】
本発明者等は、前記課題を解決すべく鋭意検討した結果、FAH12(Fatty acid hydroxylase)遺伝子が導入された酵母の形質転換体を、比較的高温で培養することにより、増殖阻害を抑制し得ること、また、高温培養により充分に培養させた後、温度シフトし、より低温で培養することによって、該形質転換体内のリシノール酸含有量を高められることを見出し、本発明を完成するに至った。
【0007】
本発明のリシノール酸産生酵母の培養方法、およびリシノール酸の製造方法は下記[1]〜[6]である。
[1]FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、
前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、
を有することを特徴とするリシノール酸産生酵母の培養方法。
[2]前記低温培養工程における前記形質転換体の培養温度が、10〜33℃である前記[1]のリシノール酸産生酵母の培養方法。
[3]前記酵母がシゾサッカロミセス(Schizosaccharomyces)属酵母である前記[1]または[2]のリシノール酸産生酵母の培養方法。
[4]前記FAH12遺伝子が、トウゴマ(Ricinus communis)、レスクェレラ・フェンドレリ(Lesquerella fendleri)または麦角菌(Claviceps purpurea)由来である前記[1]〜[3]のいずれのリシノール酸産生酵母の培養方法。
[5]前記[1]〜[4]のいずれのリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法。
【発明の効果】
【0008】
本発明のリシノール酸産生酵母の培養方法により、リシノール酸産生酵母を、増殖性とリシノール酸生産性のバランスをとりながら効率よく培養できる。
また、該形質転換体を用いた本発明のリシノール酸の製造方法により、産生されたリシノール酸を簡便に取得できる。
【図面の簡単な説明】
【0009】
【図1】pSL10−CpFAH12およびpSL10−LfFAH12のベクターの構築手順を模式的に示した図である。
【図2】pREP1−RcFAH12、pREP1−LfFAH12、およびpREP1−CpFAH12のベクターの構築手順を模式的に示した図である。
【図3】YEp352−CpFAH12のベクターの構築手順を模式的に示した図である。
【図4】参考例1において、S.ポンベおよびS.セレビシエの脂肪酸含有量を測定した結果を示した図である。
【図5】参考例3において、pREP1−LfFAH12を導入したS.ポンベの形質転換体のガスクロマトグラフ分析の結果のチャート図である。
【図6】実施例1において、各形質転換体をスポットしたプレートをインキュベートした後の各プレートの写真図である。
【図7】実施例2において、各形質転換体を、30℃または37℃で培養した場合の増殖曲線を示した図である。
【図8】実施例3における各培養条件のスキーム図である。
【図9】実施例3において、各培養条件における培養液のOD600、培養液1mL当たりの乾燥菌体重量、および培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図10】実施例3において、脂肪酸組成の経時変化を示した図である。
【図11】実施例4における各培養条件のスキーム図である。
【図12】実施例4において、各培養条件における培養液のOD600、培養液1mL当たりの乾燥菌体重量、および培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図13】実施例5における各培養条件のスキーム図である。
【図14】実施例5において、各培養液の温度シフト時点および培養終了時点の培養液のOD600、培養終了時点の培養液1mL当たりの乾燥菌体重量、並びに培養終了時点の培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図15】実施例6における各培養条件のスキーム図である。
【図16】実施例6において、各培養液の培養終了時点の培養液のOD600、および培養終了時点の培養液1mL当たりの各種脂肪酸の含有量を測定した結果を示した図である。
【図17】実施例7において、各株のガスクロマトグラフ分析の結果のチャート図である。
【図18】実施例7において、各株の培養液1mL当たりの各種脂肪酸の、総脂肪酸含有量に対する含有割合(%)の結果を示した図である。
【図19】実施例8において、各形質転換体をスポットしたプレートをインキュベートした後の各プレートの写真図である。
【図20】実施例9において、各株の培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有比率の結果を示した図である。
【図21】参考例4において、各形質転換体を、20℃または30℃で培養した場合の増殖曲線を示した図である。
【図22】実施例10において、各株の脂肪酸組成の結果を示した図である。
【発明を実施するための形態】
【0010】
[FAH12遺伝子が導入された酵母の形質転換体(リシノール酸産生酵母)]
本発明におけるリシノール酸産生酵母は、FAH12遺伝子が導入された酵母の形質転換体である。酵母に導入するFAH12遺伝子は、オレイン酸の12位の炭素原子に水酸基を導入し得るΔ12−ヒドロキシラーゼ活性を有する酵素をコードする遺伝子であれば、特に限定されず、微生物由来であってもよく、植物由来であってもよい。具体的には、例えば、トウゴマのFAH12(RcFAH12)(GenBank ID.No.: AAC49010.1)をコードする遺伝子(RcFAH12遺伝子)、レスクェレラ・フェンドレリのFAH12(LfFAH12)(GenBank ID.No.: AAC32755.1)をコードする遺伝子(LfFAH12遺伝子)、麦角菌のFAH12(CpFAH12)(GenBank ID.No.: ACF37070.1)をコードする遺伝子(CpFAH12遺伝子)、フィサリア・リンドヘイメリ(Physaria lindheimeri)のoleate 12−hydroxylase(GenBank ID.No.:ABQ01458.1)をコードする遺伝子、アスペルギルス・ニガー(Aspergillus niger)のunnamed protein product(GenBank ID.No.:CAK37451.1)をコードする遺伝子、が挙げられる。なお、GenBankはNCBI(National Center for Biotechnology Information)のデータベースである。本発明では、RcFAH12遺伝子、LfFAH12遺伝子またはCpFAH12遺伝子が好ましく、Δ12−ヒドロキシラーゼ活性がより高いため、CpFAH12遺伝子がより好ましい。
【0011】
(酵母)
FAH12遺伝子が導入される宿主は、酵母であれば特に限定されず、出芽酵母(Saccharomyces cerevisiae、以下、S.セレビシエ)をはじめとするサッカロミセス(Saccharomyces)属酵母、シゾサッカロミセス(Schizosaccharomyces)属酵母、クリュイベロミセス(Kluyveromyces)属酵母、ピキア(Pichia)属酵母、カンジダ(Candida)属酵母が挙げられる。中でも、オレイン酸含有量が高いことから、シゾサッカロミセス属酵母であることが好ましい。シゾサッカロミセス属酵母としては、たとえば、S.ポンベ、シゾサッカロミセス・ジャポニカス(Schizosaccharomyces japonicus)、シゾサッカロミセス・オクトスポラス(Schizosaccharomyces octosporus)が挙げられ、S.ポンベが好ましい。
【0012】
また、宿主とする酵母は、野生型であってもよく、用途に応じて特定の遺伝子を欠失または失活させた変異型であってもよい。特定の遺伝子を欠失または失活させる方法としては、公知の方法を用いられる。具体的には、Latour法(Nucleic Acids Research誌、第34巻、e11ページ、2006年;国際公開第2007/063919号パンフレット等に記載)を用いることにより遺伝子を欠失させられる。また、変異剤を用いた突然変異分離法(酵母分子遺伝学実験法、1996年、学会出版センター)や、PCR(ポリメラーゼ連鎖反応)を利用したランダム変異法(PCR Methods Application誌、第2巻、28-33ページ、1992年)等により遺伝子の一部に変異を導入することにより該遺伝子を失活させられる。特定遺伝子を欠失または失活させたシゾサッカロミセス属酵母宿主としては、たとえば、国際公開第2002/101038号、国際公開第2007/015470号等に記載されている。
また、特定の遺伝子の削除または不活性化を行う部分はORF(オープンリーディングフレーム)部分であってもよく、発現調節配列部分であってもよい。特に好ましい方法は、構造遺伝子のORF部分をマーカー遺伝子に置換するPCR媒介相同組換え法(Yeast誌、第14巻、943-951ページ、1998年)による削除または不活性化の方法である。
【0013】
さらに宿主となる酵母には、形質転換体を選択するためのマーカーを有するものを用いることが好ましい。たとえば、ある遺伝子が欠落していることにより特定の栄養成分が生育に必須である宿主を使用することが好ましい。目的遺伝子配列を含むベクターにより形質転換をして形質転換体を作製する場合、ベクターにこの欠落している遺伝子(栄養要求性相補マーカー)を組み込んでおくことにより、形質転換体は宿主の栄養要求性が消失する。この宿主と形質転換体の栄養要求性の相違により、両者を区別して形質転換体を得られる。
たとえば、オロチジン5’−リン酸デカルボキシラーゼ遺伝子(ura4遺伝子)が欠失または失活してウラシル要求性となっている酵母を宿主とし、ura4遺伝子(栄養要求性相補マーカー)を有するベクターにより形質転換した後、ウラシル要求性が消失したものを選択することにより、ベクターが組み込まれた形質転換体を得られる。宿主において欠落により栄養要求性となる遺伝子は、形質転換体の選択に用いられるものであればura4遺伝子には限定されず、イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leu1遺伝子)等であってもよい。
【0014】
(発現カセット)
本発明におけるリシノール酸産生酵母は、FAH12遺伝子をコードする領域を含む構造遺伝子配列(FAH12遺伝子配列)、並びに該遺伝子を発現させるためのプロモーター配列およびターミネーター配列を含む発現カセットを、酵母に導入することにより作製できる。FAH12遺伝子配列としては、麦角菌等由来のFAH12をコードする遺伝子をそのまま用いてもよいが、宿主として用いる酵母内での発現量を増大させるために、前記遺伝子配列を、宿主での高発現遺伝子において使用頻度の高いコドンに改変することが好ましい。
【0015】
発現カセットとは、蛋白質を発現するために必要なDNAの組み合わせであり、該蛋白質をコードする構造遺伝子と宿主である酵母内で機能するプロモーターとターミネーターとを含む。
前記発現カセットには、さらに、5’−非翻訳領域、3’−非翻訳領域のいずれか1つ以上が含まれていてもよい。好ましい発現カセットは、FAH12遺伝子配列、プロモーター、ターミネーター、5’−非翻訳領域、3’−非翻訳領域を全て含む発現カセットである。さらに、栄養要求性相補マーカー等の遺伝子を有していてもよい。
【0016】
プロモーターとターミネーターは、宿主である酵母内で機能してFAH12を発現できるものであればよい。酵母内で機能するプロモーターとしては、酵母が本来有するプロモーター(転写開始活性が高いものが好ましい)や酵母が本来有しないプロモーター(ウイルス由来のプロモーターなど)を使用できる。プロモーターはベクター内に2種以上存在していてもよい。
シゾサッカロミセス属酵母が本来有するプロモーターとしては、たとえば、アルコールデヒドロゲナーゼ遺伝子プロモーター、チアミンの代謝に関与するnmt1遺伝子プロモーター、グルコースの代謝に関与するフルクトース−1、6−ビスホスファターゼ遺伝子プロモーター、カタボライト抑制に関与するインベルターゼ遺伝子のプロモーター(国際公開第99/23223号参照)、熱ショック蛋白質遺伝子プロモーター(国際公開第2007/26617号参照)などが挙げられる。
シゾサッカロミセス属酵母が本来有しないプロモーターとしては、たとえば、特開平5−15380号公報、特開平7−163373号公報、特開平10−234375号公報に記載されている動物細胞ウイルス由来のプロモーターが挙げられ、hCMVプロモーター、SV40プロモーターが好ましい。
シゾサッカロミセス属酵母内で機能するターミネーターとしては、シゾサッカロミセス属酵母が本来有するターミネーターやシゾサッカロミセス属酵母が本来有しないターミネーターを使用できる。ターミネーターはベクター内に2種以上存在していてもよい。
ターミネーターとしては、たとえば、特開平5−15380号公報、特開平7−163373号公報、特開平10−234375号公報に記載されているヒト由来のターミネーターが挙げられ、ヒトリポコルチンIのターミネーターが好ましい。
【0017】
(ベクター)
本発明におけるリシノール酸産生酵母は、具体的には、前記発現カセットを含むベクターにより宿主である酵母を形質転換することにより得られる。形質転換に用いるベクターは、環状DNA構造または線状DNA構造を有するベクターに、該発現カセットを組み込むことにより製造できる。該発現カセットが、宿主の細胞内で染色体外遺伝子として保持される形質転換体を作製する場合には、該ベクターは、宿主細胞内で複製されるための配列、即ち、自律複製配列(Autonomously Replicating Sequence: ARS)を含むプラスミドであることが好ましい。一方で、該発現カセットが、宿主細胞の染色体中に組み込まれた形質転換体を作製する場合には、該ベクターは、線状DNA構造であり、かつARSを有していないものとして、宿主細胞へ導入されることが好ましい。たとえば、該ベクターは、線状DNAからなるベクターであってもよく、宿主への導入時に、線状DNAに切り開くための制限酵素部位を備える環状DNA構造のベクターであってもよい。該ベクターがARSを有するプラスミドの場合、ARS部分を削除して線状DNA構造、またはARS部分を開裂させることによりARSの機能を失活させた線状DNA構造とした後、宿主へ導入できる。
【0018】
該ベクターは、形質転換体を選択するためのマーカーを有することが好ましい。該マーカーとしては、たとえば、ura4遺伝子(栄養要求性相補マーカー)、イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leu1遺伝子)が挙げられる。
【0019】
リシノール酸産生酵母の製造に用いられる前記発現カセットを含むベクターは、たとえば、外来構造遺伝子を導入するためのクローニングサイトを備える公知の発現ベクター中の該クローニングサイトに、FAH12遺伝子配列を挿入することによって製造できる。また、公知の発現ベクターに、FAH12遺伝子配列を含む発現カセットを組み込むことによっても製造できる。
【0020】
(形質転換体の製造方法)
本発明におけるリシノール酸産生酵母は、前記発現カセットを、染色体中に有するか、または、染色体外遺伝子として有する. 発現カセットを染色体中に有するとは、宿主細胞の染色体中の1カ所以上に発現カセットが組み込まれていることであり、染色体外遺伝子として有するとは、発現カセットを含むプラスミドを細胞内に有するということである。形質転換体の継代培養が容易であることから、該発現カセットを染色体中に有することが好ましい。
【0021】
形質転換方法は、公知の形質転換方法をいずれも用いられる。該形質転換方法としては、たとえば、酢酸リチウム法、エレクトロポレーション法、スフェロプラスト法、ガラスビーズ法など従来周知の方法や、特開2005−198612号公報記載の方法が挙げられる。また、市販の酵母形質転換用キットを用いてもよい。
【0022】
形質転換を行った後、通常は得られた形質転換体を選抜する。選抜方法としては、たとえば、以下に示す方法が挙げられる。前記栄養要求性マーカーにより形質転換体を選択できる培地によりスクリーニングし、得られたコロニーから複数を選択する。選抜した形質転換体に対してパルスフィールドゲル電気泳動法によるゲノム解析を行うことにより、染色体に組み込まれたベクターの数や発現カセットの数を調べられる。
【0023】
[リシノール酸産生酵母の培養方法]
酵母にFAH12を発現させた場合、通常の培養温度(30℃程度)では増殖阻害がかかるが、比較的高温で培養した場合には増殖阻害がかからない。高温条件下では、FAH12の発現が抑制されるか、または発現したFAH12のΔ12−ヒドロキシラーゼ活性が失活若しくは低下するためと推察される。本発明では、リシノール酸産生酵母をまず高温で培養した後、培養温度を低温にシフトさせることによって、該酵母内でリシノールを産生させる。はじめに高温で培養することにより、温度シフト後にもFAH12による増殖阻害の影響を抑制しつつ、FAH12によるリシノール酸産生を促進し得る。
すなわち、本発明のリシノール酸産生酵母の培養方法は、FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程とを有することを特徴とする。
【0024】
酵母の培養温度があまりに高い場合には、温度ストレスにより増殖がかえって低下する。リシノール酸産生酵母を35℃以上40℃未満で培養することにより、過度の高温ストレスを受けることなく、FAH12による増殖阻害作用を充分に抑制し得る。高温培養工程における培養温度は、35〜39℃が好ましく、35〜38℃がより好ましい。
【0025】
低温培養工程における培養温度を、高温培養工程よりも低い温度、すなわち35℃未満にすることで、増殖阻害作用が抑制されつつ、FAH12によるリシノール酸産生が促進される。但し、酵母の培養温度があまりに低い場合には、温度ストレスにより増殖がかえって低下する。そこで、低温培養工程における培養温度は、10〜33℃が好ましく、15〜33℃がより好ましく、20〜30℃がさらに好ましい。
【0026】
低温培養工程移行後にも、リシノール酸産生酵母が充分に増殖でき、培養液当たりのリシノール酸産生量を改善し得るため、酵母の増殖がある程度盛んな状態、特に対数増殖期にある段階で、高温培養工程から低温培養工程へ移行することが好ましい。このため、本発明においては、形質転換体の培養液のOD600が5.5以下の時点で、好ましくは5以下の時点で、より好ましくは4,4以下の時点で、高温培養工程から低温培養工程へ移行する。また、高温培養工程から低温培養工程へ移行する時点における形質転換体の培養液のOD600は、0.5以上であり、0.55以上であることがより好ましく、1.4以上であることがさらに好ましい。特に、リシノール酸産生酵母を、35℃以上40℃未満で、培養液のOD600が1.4〜4.4になるまで培養した後、当該培養液の温度を低温にシフトさせてさらに培養を続けることにより、より大量のリシノール酸を産生できる。培養液及びその希釈液のOD600は、例えば、分光光度計を使用して常法により測定できる。
【0027】
本発明のリシノール酸産生酵母の培養方法では、高温培養工程および低温培養工程のいずれでも、リシノール酸産生酵母は、培養温度以外は一般的な酵母の培養方法と同様にして培養できる。たとえば、培養液には、宿主とした酵母の培養に用いられる公知の培養培地を用いることができ、宿主が資化しうる炭素源、窒素源、無機塩類等を含有し、宿主の培養を効率良く行えるものであればよい。培養液としては、天然培地を用いてもよく、合成培地を用いてもよい。
【0028】
炭素源としては、たとえばグルコース、フルクトース、スクロース等の糖が挙げられる。
窒素源としては、たとえばアンモニア、塩化アンモニウム、酢酸アンモニウム等の無機酸または無機酸のアンモニウム塩、ペプトン、カザミノ酸が挙げられる。
無機塩類としては、たとえばリン酸マグネシウム、硫酸マグネシウム、塩化ナトリウムが挙げられる。
【0029】
培養には公知の細胞培養方法を用いることができ、たとえば振盪培養、攪拌培養等により行える。
培養は、回分培養であってもよく、連続培養であってもよい。また、培養時間は適宜決定できる。
【0030】
[リシノール酸の製造方法]
本発明のリシノール酸の製造方法は、本発明のリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とする。
培養されたリシノール酸産生酵母からのリシノール酸の抽出・精製は、溶剤抽出法等の一般的に脂肪酸の抽出・精製に使用される公知の手法により実施できる。例えば、リシノール酸産生酵母を培養した後、得られた培養液を遠心分離して菌体を回収し、該菌体をメタノール中でグラスビーズ等を用いて破砕しながら、クロロホルム/メタノール(1:2)を加えて抽出し、その後精製することにより、リシノール酸を精製できる。
また、リシノール酸は、ガスクロマトグラフィ法等の公知の手法で検出および定量できる。
【実施例】
【0031】
以下、実施例等を示して本発明を詳細に説明する。ただし、本発明は以下の記載によっては限定されない。
なお、以下の実施例等において、酵母の培養液および該希釈液のOD値は、分光光度計UV1600(島津製作所製)を用いて測定した。
【0032】
[酵母]
以下の実施例等において、S.ポンベは、ロイシンおよびウラシル要求株ARC010(遺伝子型:h− 、leu1−32、ura4−D18)(国際公開第2007/015470号パンフレット参照。)を用いた。また、S.セレビシエは、BY4741(遺伝子型:MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0)を用いた。
S.ポンベは、プラスミドを維持するために必要な選択圧に依存して、EMMSまたはEMMSドロップアウト培地中で培養した(Alfa et al., 1993 ,“A laboratory course manual.”, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.)。EMMSは通常の培養時に使用され、窒素が制限された最小培地(EMM−C/N3)がリシノール酸産生には使用された。発明者らによる従前の実験結果から、EMM−C/N3中で培養したほうが脂肪酸含有量が高くなることが示唆されていたためである。EMM培地は、0.5%(w/v)の塩化アンモニウム、2%(w/v)のグルコースを含有していた。一方、EMM−C/N3は、塩化アンモニウムの濃度は0.1%にまで低減されており、グルコース濃度が10%にまで増大していた。nmt1プロモーターの下、遺伝子発現を抑制するために、15μM(5μg/mL)のチアミンを使用した。
S.セレビシエは、プラスミドを維持するために必要な選択圧に依存して、最小合成培地(SD)、合成完全培地(SC)または合成完全ドロップアウト培地中で培養した(Sherman, et.al., 1986, “Methods in yeast genetics. ”, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.)。最小合成培地またはSCは、0.17%(w/v)のアミノ酸およびアンモニウム塩不含有イーストニトロジェンベース(Bactoyeast nitrogen base、Difco社製)、2%(w/v)のグルコース、および0.5%(w/v)の硫酸アンモニウムを含有していた。窒素が制限された最小培地(NSD)では、硫酸アンモニウム含有量が0.1%にまで低減されており、グルコース濃度が10%にまで増大していた(Yazawa, et.al., 2007,“Heterologous production of dihomo-gamma-linolenic acid in yeast Saccharomyces cerevisiae.”, Appl Env Microbiol, vol.73, pp.6965-6971)。
【0033】
[プラスミド構築]
RcFAH12遺伝子、LfFAH12遺伝子、およびCpFAH12遺伝子のコード領域を含む酵母の形質転換用プラスミドを構築した。DNA操作の標準的な技術は、SambrookおよびRussellの方法(“Molecular cloning: A laboratory manual. 3rd ed.”, 2001, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.)に準じて行った。また、プラスミド構築は、大腸菌DH5α(F−endA1、hsdR17、supE44、thi−1、recA1、gyrA96、relA1、Δ(argF−lacZ)U169、Φ80、Δ(lacZ)M15)株を用いて行った。当該DH5α株は、アンピシリンまたはカナマイシンを添加したLB培地(Luria-Bertani broth)中で培養した。
【0034】
CpFAH12遺伝子のDNA配列(Meesapyodsuk and Qiu, 2008, Plant Physiology, vol.147, pp.1325−1333)は、S.ポンベにおいて好ましい使用頻度のコドンに改変した後、GENEART社(Regensburg, Germany)に依頼して化学合成した。合成されたCpFAH12のORFは、5’末端と3’末端にそれぞれ人工的に付加されていた制限酵素部位を使用して、大腸菌のプラスミドpMK−CpFAH12から、制限酵素NcoIおよびSalIで二重消化したフラグメントとして切り出し、pL2428−9(略してpSL10−CpFAH12)を構築するために、マルチクローニングサイトの制限酵素AarIおよびSalIで二重消化したプラスミドpSL10に組み込んだ。合成されたプラスミドをpSL10−CpFAH12とした。pSL10は、leu1マーカーを有する、染色体へ組み込まれるタイプのプラスミドであり、pSL6プラスミド(Alimjan et al., Appl Microbiol Biotechnol, 2010, vol.85, pp.667−677)のhCMVプロモーター(hCMV−p)をnmt1プロモーターに置換し、LPIターミネーター(LPI−ter.)をinv1ターミネーターに置換したプラスミドである。pSL10の塩基配列を配列番号1に示す。S.ポンベにpSL10−CpFAH12を導入した形質転換体中では、チアミン非存在時には、nmt1プロモーターの制御下でCpFAH12が発現したが、チアミン存在時には発現は抑制された。
pSL10−CpFAH12(pL2428−9)と同様にして、LfFAH12遺伝子のORFを組み込んだpL2427−8 (略してpSL10−LfFAH12)を構築した。
図1に、pSL10−CpFAH12およびpSL10−LfFAH12のベクターの構築手順を模式的に示した。なお、同様な手法によりpSL10−RcFAH12も作製できる。
【0035】
また、pMK−CpFAH12から、制限酵素HindIIIおよびPvuIIで二重消化したフラグメントとしてCpFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。合成されたプラスミドをpL2414−1(略してpREP1−CpFAH12)とした。pREP1は、S.セレビシエのLEU2マーカーを有する染色体外に存在するタイプのプラスミドである。S.ポンベにpREP1−CpFAH12を導入した形質転換体中では、チアミン非存在時には、nmt1プロモーターの制御下でCpFAH12が発現したが、チアミン存在時には発現は抑制された。
pL2390−1(略してpREP1−RcFAH12)は、pGA4−RcFAH12H6を制限酵素NcoIおよびEcl136IIで二重消化したフラグメントとしてRcFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。pL2391−63(略してpREP1−LfFAH12)は、pMA−LfFAH12を制限酵素SmaIおよびPvuIIで二重消化したフラグメントとしてLfFAH12を切り出し、マルチクローニングサイトの制限酵素NdeIで消化したプラスミドpREP1に組み込んだ。
図2に、pREP1−RcFAH12、pREP1−LfFAH12、およびpREP1−CpFAH12のベクターの構築手順を模式的に示した。
【0036】
また、pMK−CpFAH12から、制限酵素EcoRIおよびXhoIで二重消化したフラグメントとしてCpFAH12を切り出し、pL2379−2を構築するために、マルチクローニングサイトの制限酵素EcoRIおよびXhoIで二重消化したプラスミドYEp352GAPに組み込んだ。S.セレビシエにpL2379−2(略してYEp352−CpFAH12)を導入した形質転換体中では、解糖系のGAPDHプロモーターの制御下でCpFAH12が発現した。
図3に、YEp352−CpFAH12のベクターの構築手順を模式的に示した。なお、同様な手法によりYEp352−RcFAH12も作製できる。
【0037】
[形質転換体の作製]
酵母細胞は、Frozen−EZ Yeast Transformation II kit (Zymo Research, CA, USA)を用いて、製造者により推奨されるプロトコールに従って形質転換した。
たとえば、CpFAH12遺伝子を染色体中に組み込む形で導入したS.ポンベの形質転換体の作製は、以下のようにして行った。すなわち、酵母の第2染色体のleu1座にpSL10−CpFAH12を組み込むため、pSL10−CpFAH12は制限酵素BsiWIによって消化し、得られた線状プラスミドをS.ポンベARC010−1株に導入し、leu1+の形質転換体を選抜した。
一方で、CpFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体は、pREP1−CpFAH12をそのままS.ポンベARC010−1株に導入し、LEU2+の形質転換体(すなわち、ロイシン要求性のない形質転換体)を選抜することによって作製した。
RcFAH12遺伝子またはLfFAH12遺伝子を染色体中または染色体外に導入したS.ポンベの形質転換体は、CpFAH12遺伝子を染色体中または染色体外に導入したS.ポンベの形質転換体と同様にして作製した。
【0038】
[参考例1]
S.ポンベおよびS.セレビシエの脂肪酸含有量を測定した。
具体的には、S.ポンベの場合は、EMM−C/N3(ロイシンおよび/またはウラシル無含有培地)中で、30℃で一晩前培養し、該前培養物を新しいEMM−C/N3に接種し、40rpmで振とう培養した。S.セレビシエの場合は、NSD培地を用いた。
得られた培養物の総脂肪酸含有量は、ガスクロマトグラフ分析によるKainouらの方法(2006, Yeast, vol.23, pp.605-612)に準じて行った。具体的には、以下に示す通りである。まず、脂肪酸分析は、温度制御環境下(160〜234℃、昇温速度:4.5℃/min)で、TC−70キャピラリーカラム(30m×0.25mm(内径)、GL Sciences製)を備えたガスクロマトグラフ(GC2010、島津製作所製)に0.2μLの培養液を投与して実施した。脂肪酸組成は、各ピークの面積に基づいて算出され、含有量は、標準としたヘプタデカン酸メチル(C17:0)と比較することによって決定した。
【0039】
測定結果を図4に示す。縦軸は全脂肪酸に対する各脂肪酸の含有量割合(%)である。S.セレビシエでは、オレイン酸(C18:1)は30%強含まれていたが、パルミトレイン酸(C16:1)が約45%と最も多かった。一方、S.ポンベでは、80%強がオレイン酸(C18:1)であり、その他の脂肪酸含有量はいずれも非常に少なかった。
【0040】
[参考例2]
pREP1、pREP1−CpFAH12、pREP1−RcFAH12、またはpREP1−LfFAH12を導入したS.ポンベの形質転換体を、それぞれチアミン存在下および非存在下で培養し、増殖能を調べた。
具体的には、まず、チアミン含有EMM−C/N3−LEU(EMM−C/N3からロイシンが除去された培地)中、30℃で一晩前培養した前培養液を、チアミン不添加のEMM−C/N3−LEUのプレート(アガー平板培地)またはチアミン含有EMM−C/N3−LEUのプレートに塗布し、30℃で4日間培養し、コロニーの形成の有無や、形成されたコロニーのサイズを調べた。
空ベクターであるpREP1を導入した形質転換体をチアミン含有プレートに塗布した場合に形成されたコロニーのサイズを基準として、その他の形質転換体の増殖能を評価した。評価結果を表1に示す。表1中、「+++」は生育が正常であり、「++」は生育が若干悪く、「+」は生育が悪く、「−」は増殖できなかったことを意味する。また、「遺伝子発現OFF(チアミン有り)」はチアミン含有EMM−C/N3−LEUプレートの結果を、「遺伝子発現ON(チアミン無し)」はチアミン不添加のEMM−C/N3−LEUプレートの結果を、それぞれ示す。
【0041】
【表1】
【0042】
RcFAH12を発現させた場合には、外来蛋白質を発現させていない場合と同程度の増殖能であったが、LfFAH12を発現させた形質転換体は、RcFAH12を発現させた形質転換体よりも増殖能が低下しており、CpFAH12を発現させた形質転換体は非常に増殖能が低下していることがわかった。つまり、LfFAH12やCpFAH12を発現させることによって、増殖能が低下した。
【0043】
[参考例3]
pREP1−LfFAH12を導入したS.ポンベの形質転換体を培養し、リシノール酸をはじめとする脂肪酸の含有量を調べた。
具体的には、まず、チアミン含有EMM−C/N3−LEU中、30℃で一晩前培養した前培養液を、EMM−C/N3−LEU中に接種し、30℃、40rpmで5日間振とう培養した。得られた培養物の各脂肪酸の含有量を、参考例1と同様にしてガスクロマトグラフ分析によって測定した。測定の結果得られたチャート図を図5に示す。該形質転換体では、リシノール酸の標準サンプルと同じ位置にピークが検出された。すなわち、該形質転換体ではリシノール酸が生産されていることが確認できた。
【0044】
[実施例1]
CpFAH12遺伝子を染色体中に組み込んだS.ポンベの形質転換体(以下、CpFAH12インテグラント株)の各培養温度における増殖能を調べた。
具体的には、CpFAH12インテグラント株にura4マーカーを含む空ベクターpREP2を導入した形質転換体(CpFAH12株)と、S.ポンベにpSL10およびpREP2を導入した形質転換体(コントロール株)を、それぞれチアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一晩、培養液のOD600が約4になるまで前培養した。該前培養液を同種の新しい培地で希釈し、さらに、10倍希釈系列を調整し、各10μLずつを、15μMのチアミン含有EMM−C/N3−URA,LEUプレート(発現抑制条件)またはチアミン不添加のEMM−C/N3−URA,LEUプレート(発現誘導条件)上にスポットした。該プレートを37℃、35℃、30℃、25℃、または20℃で4〜9日間インキュベートした。
インキュベート後の各プレート表面の写真図を図6に示す。図中、「Control」はコントロール株の結果であり、「CpFAH12」はCpFAH12株の結果を示す。また、図中、白い部分がコロニーである。チアミン含有プレート(図中、「+Thiamine」)では、20℃、9日間培養した場合のみ、CpFAH12株がコントロール株よりもやや増殖能が低かったものの、その他の培養条件では両形質転換体の増殖能に特段の差はなかった。一方、参考例2ではCpFAH12を発現させた株ではほとんど増殖が観察されなかったにも関わらず、チアミン不含有プレート(図中、「No Thiamine」)では、濃い濃度でスポットとした場合に、全ての培養条件でCpFAH12株の増殖が確認できた。但し、37℃、4日間培養した場合を除き、CpFAH12株のほうがコントロール株よりも明らかに増殖能が低かった。すなわち、CpFAH12の発現誘導条件下のCpFAH12インテグラントの増殖能は培養温度に影響され、37℃で前培養した後に培養温度を低下させた場合には、増殖能は低下するものの、増殖自体は可能であること、および37℃ではほとんど低下しないことがわかった。
【0045】
[実施例2]
実施例1で用いたCpFAH12株およびコントロール株の、30℃または37℃の液体培地中の増殖能を調べた。
具体的には、CpFAH12株およびコントロール株を、チアミン不添加のEMM−C/N3−URA,LEU中、37℃で一晩前培養した。該前培養物を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、30℃または37℃で36時間、攪拌培養した。630nmの濁度を自動検出器(Bio−Plotter、東洋測器製)でモニターすることにより、液体培地中の増殖曲線を描いた。図7に増殖曲線を示す。図7(A)は30℃で培養した結果であり、図7(B)は37℃で培養した結果である。図中、三角がCpFAH12株の増殖曲線を、円がコントロール株の増殖曲線を、それぞれ示す。37℃で培養した場合よりも、30℃で培養した場合には、CpFAH12株はコントロール株よりも明らかに増殖が抑制されていた。
【0046】
[実施例3]
37℃から20℃または15℃に培養温度をシフトさせるタイミングの、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
図8は、評価方法のスキーム図である。まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、最初に37℃で1日間(OD600が5、対数増殖期後期)または2日間(OD600が6、定常増殖期初期)培養し、その後20℃または15℃に培養温度をシフトさせて5日間培養した。対照として、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、温度シフトせずに37℃で7日間培養した(図8の下段)。液体培養は全て、40rpmで振とう培養した。
【0047】
本培養開始後1日経過ごとに、培養液の一部をサンプリングし、培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液のOD600は実施例2と同様にして、各種脂肪酸の含有量は参考例1と同様にして、それぞれ測定した。培養液のOD600および培養液1mL当たりの乾燥菌体重量の測定結果を図9(A)に、各種脂肪酸の含有量の測定結果を図9(B)に、それぞれ示す。図9(B)中、「C18:1−OH」はリシノール酸を、「C18:2」はリノレン酸を、「Others」はその他の脂肪酸の総和を、それぞれ示す。培養液のOD600および培養液1mL当たりの乾燥菌体重量は、培養温度の温度シフトによってはあまり差がなかった。しかし、各種脂肪酸の含有量は、温度シフトせずに37℃で培養し続けた場合、37℃で2日間培養した後に20℃または15℃にシフトした場合には、リシノール酸の含有量は非常に低かった。これに対して、37℃で1日間培養した後に20℃または15℃にシフトした場合には、培養時間の経過とともにリシノール酸の含有量が顕著に増大していた。該結果から、培養温度が37℃の場合、および増殖が定常状態になった後に低温へ温度シフトした場合には、リシノール酸が産生されないことが分かった。培養温度が37℃の場合等でリシノール酸産生が行われないのは、CpFAH12が発現しないため、またはCpFAH12の活性が非常に低下するためと推察される。
【0048】
さらに、図10に、脂肪酸組成の経時的変化を示す。図10(A)は37℃で1日間培養した後に20℃にシフトして5日間培養した場合、図10(B)は37℃で2日間培養した後に20℃にシフトして5日間培養した場合、図10(C)は37℃で7日間培養した場合の結果である。37℃で2日間培養した後に20℃にシフトした場合には、37℃で7日間培養した場合と同様、脂肪酸組成はほとんど経時的変化がなかった。これに対して37℃で1日間培養した後に20℃にシフトした場合には、培養時間の経過と共にオレイン酸含有割合が低下し、逆にリシノール酸含有割合が増大した。その他の脂肪酸の含有割合は、ほとんど経時的変化がなかった。
【0049】
[実施例4]
37℃から20℃、25℃または30℃への培養温度シフトの、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図11に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、20℃、25℃または30℃で10日間、40rpmで振とう培養した(図11の上段)。対照として、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、温度シフトせずに37℃で10日間、40rpmで振とう培養した(図11の下段)。
実施例3と同様に、本培養開始後1日経過ごとに、培養液の一部をサンプリングし、培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液のOD600および培養液1mL当たりの乾燥菌体重量の測定結果を図12(A)に、各種脂肪酸の含有量の測定結果を図12(B)に、それぞれ示す。図12(B)中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。
【0050】
最終的なリシノール酸産生量は、本培養を20℃で実施した場合が、25℃または30℃で実施した場合よりも多くなることが確認された。しかし、本培養を25℃で行うほうが、利点があると考えられる。細胞増殖は25℃のほうが良好であるため、本培養開始4日目では、25℃の培養条件のほうが20℃よりもリシノール酸の産生量が高かったためである。具体的には、培養液当たりのリシノール酸産生量は、20℃で10日間培養後には163.2μg/mLであったが、20℃で4日間培養後には48.2μg/mLでしかなかった。これに対して25℃で4日間培養後には108.8μg/mLであり、該生産量は、20℃で10日間培養した場合の66%であった。さらにコストの点から、冷却コストを低減でき、かつ培養日数を10日間から4日間に短縮することで培養コストを低減できるため、細胞増殖は20℃よりも25℃で行うほうが好ましい。このように、冷却コストを低減し、培養時間を短縮することで製造コストを低減できるため、工業上の観点からは、25℃培養のほうが20℃培養よりも好ましい。
【0051】
[実施例5]
37℃から20℃に培養温度をシフトさせる時点の酵母の増殖状態の、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図13に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.01〜0.16の間様々な値になるように希釈した5つの本培養液を調製後、37℃で1日間、40rpmで振とう培養した後、培養液の温度を20℃にシフトし、シフト後さらに5日間、40rpmで振とう培養した(図13の上段)。5つの培養液の温度シフト時のOD600は、それぞれ、0.55(培養液A)、1.4(培養液B)、2.5(培養液C)、4.4(培養液D)および5.5(培養液E)であった。対照として、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、温度シフトせずに37℃で6日間、40rpmで振とう培養した(図13の下段)。
実施例3と同様にして、本培養終了時点の培養液のOD600、培養液1mL当たりの乾燥菌体重量(DCW)、および培養液1mL当たりの各種脂肪酸の含有量を測定した。温度シフト開始時点および培養終了時点の培養液のOD600、並びに培養終了時点の培養液1mL当たりの乾燥菌体重量の測定結果を図14(A)に、培養終了時点の培養液1mL当たりの各種脂肪酸の含有量の測定結果を図14(B)に、それぞれ示す。図14(B)中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。さらに、総脂肪酸含有量に対する各脂肪酸の含有量比(質量%)を表2に示す。図14および表2の各測定値は、独立した2回の試行を行い、その平均値である。
【0052】
【表2】
【0053】
図14(B)および表2に示す通り、培養液A〜Eの全てでリシノール酸の産生が確認されたが、温度シフト時点の培養液のOD600が1.4〜4.4である培養液B〜Dが、培養液AおよびEよりもリシノール酸産生量が多かった。該結果から、酵母が対数増殖期にある時点で温度シフトを行うことにより、より大量のリシノール酸を産生させ得ることが示唆された。
【0054】
[実施例6]
37℃から20℃に培養温度をシフトさせる時点の培養液の濃度の、CpFAH12インテグラント株のリシノール酸産生に対する影響を評価した。
具体的には、図15に示すように、まず、CpFAH12インテグラント株を、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.07になるように希釈した後、37℃で培養液のOD600が2.5になるまで40rpmで振とう培養した後、培養液を遠心分離処理して菌体を回収した後、遠心分離処理前の菌体濃度の1倍、2倍、または4倍となるように、元々の培地または新しい同種の培地に懸濁して培養液を調製した。該培養液の温度を20℃にシフトし、シフト後さらに5日間、40rpmで振とう培養した。
実施例3と同様にして、本培養終了時点の培養液のOD600、および培養液1mL当たりの各種脂肪酸の含有量を測定した。測定結果を図16に示す。図16中、「C18:1−OH」、「C18:2」および「Others」は図9(B)と同様である。また、「1×Used」、「2×Used」および「4×Used」は、元々の培地で菌体濃度が遠心分離処理前の菌体濃度の1倍、2倍、または4倍になるように調製した培養液の結果であり、「1×Fresh」、「2×Fresh」および「4×Fresh」は、新しい培地で菌体濃度が遠心分離処理前の菌体濃度の1倍、2倍、または4倍になるように調製した培養液の結果である。
【0055】
遠心分離処理後の培養液の調製に新しい培地を用いたほうが、元々の培地を用いるよりも細胞増殖速度が速く、よって培養液1mL当たりのリシノール酸産生量も多くなった。また、遠心分離処理後の菌体の濃縮倍率が大きいほど、培養液1mL当たりのリシノール酸産生量も多くなった。しかし、総脂肪酸全体に占めるリシノール酸の含有割合は、各培養液において特に大きな差はなかった。
【0056】
[実施例7]
37℃から20℃または30℃への培養温度シフトの、実施例1で用いたCpFAH12株およびコントロール株のリシノール酸産生に対する影響を評価した。
まず、CpFAH12株およびコントロール株をそれぞれ、チアミンが添加されていないEMM−C/N3−URA,LEU中、37℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.5になるように希釈した後、20℃、30℃または37℃に培養温度をシフトさせて4日間培養した。本培養開始後1日経過ごとに、培養液の一部をサンプリングし、参考例1と同様にして、培養液1mL当たりの各種脂肪酸の含有量を測定した。
本培養を20℃で行った場合の各株のガスクロマトグラフ分析の結果のチャート図を図17に示す。CpFAH12株では、リシノール酸の標準サンプルと同じ位置にピークが検出された。該ピークは、CpFAH12を発現していないコントロール株では観察されなかった。
培養液1mL当たりの各種脂肪酸の、総脂肪酸含有量に対する含有割合(%)の結果を図18に示す。図18(A)はコントロール株の結果であり、図18(B)はCpFAH12株の結果である。CpFAH12を発現していないコントロール株では、本培養の温度に関わらず、脂肪酸組成は全てほぼ同様であった。これに対してCpFAH12株では、37℃で培養した場合にはコントロール株の結果とほぼ変わらなかったのに対して、20℃および30℃で培養した場合には、オレイン酸の含有割合の低下に伴い、リシノール酸の含有割合が増大していた。20℃の場合には、培養日数が長くなるにつれてリシノール酸含有割合が増大していた。30℃の場合には、培養日数が2日目でリシノール酸の含有割合はピークを迎えていた。
【0057】
[実施例8]
pREP1、pREP1−CpFAH12、pREP1−RcFAH12、またはpREP1−LfFAH12を導入したS.ポンベの形質転換体を、それぞれチアミン存在下および非存在下で培養し、各培養温度における増殖能を調べた。
まず、各形質転換体をそれぞれ、チアミンが添加されていないEMM−C/N3−LEU中、37℃で一日間前培養した。次いで、該前培養液を同種の新しい培地で希釈し、さらに、10倍希釈系列を調整し、各10μLずつを、15μMのチアミン含有EMM−C/N3−LEUプレート(発現抑制条件)またはチアミン不添加のEMM−C/N3−LEUプレート(発現誘導条件)上にスポットした。該プレートを20℃で10日間、30℃で5日間、または37℃で5日間インキュベートした。インキュベート後の各プレート表面の写真図を図19に示す。図中、白い部分がコロニーである。図中、「+Thiamine」がチアミン含有プレートであり、「No Thiamine」がチアミン不含有プレートである。
チアミン不含有プレートでは、培養温度にかかわらず、pREP1−CpFAH12を導入した形質転換体およびpREP1−LfFAH12を導入した形質転換体では、pREP1を導入した形質転換体よりも増殖能が低い傾向が観察された。
【0058】
[実施例9]
RcFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体(以下、pREP1−RcFAH12導入株)およびLfFAH12遺伝子を染色体外遺伝子として導入したS.ポンベの形質転換体(以下、pREP1−LfFAH12導入株)の30℃で培養した場合の脂肪酸組成を調べた。対照として、pREP1を導入したS.ポンベの形質転換体(以下、pREP1導入株)を30℃で培養した場合の脂肪酸組成を調べた。
まず、各株をチアミンが添加されていないEMM−C/N3−LEU中、30℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地またはチアミン含有EMM−C/N3−LEUで、OD600が0.05になるように希釈した後、30℃で7日間、40rpmで振とう培養した。参考例1と同様にして、培養終了後の培養液1mL当たりの各種脂肪酸の含有量を測定した。培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有比率の結果を図20に示す。図20中、「pREP1」、「RcFAH12」、および「LfFAH12」はそれぞれ、pREP1導入株、pREP1−RcFAH12導入株、およびpREP1−LfFAH12導入株をチアミンが添加されていないEMM−C/N3−LEU中で本培養した結果を示し、「pREP1+Thi」、「RcFAH12+Thi」、および「LfFAH12+Thi」はそれぞれ、pREP1導入株、pREP1−RcFAH12導入株、およびpREP1−LfFAH12導入株をチアミン含有EMM−C/N3−LEU中で本培養した結果を示す。
【0059】
図20に示すように、pREP1−RcFAH12導入株およびpREP1−LfFAH12導入株では、チアミン含有培地ではpREP1導入株と同様、リシノール酸はほとんど含有されていなかったが、チアミンが添加されていない培地では、オレイン酸含有割合が低下し、リシノール酸の含有割合が上昇していた。すなわち、CpFAH12遺伝子を導入した形質転換体と同様、RcFAH12遺伝子またはLfFAH12遺伝子を導入した形質転換体では、30℃で培養した場合に、リシノール酸が産生されることがわかった。
【0060】
[参考例4]
CpFAH12遺伝子を染色体外遺伝子として導入したS.セレビシエの形質転換体の増殖能を調べた。
具体的には、該S.セレビシエにYEp352−CpFAH12を導入した形質転換体(CpFAH12株)と、S.セレビシエに空ベクターYEp352GAPを導入した形質転換体(コントロール株)を、それぞれSC−URA中、30℃で一晩前培養した。該前培養物を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、20℃または30℃で攪拌培養した。630nmの濁度を自動検出器(Bio−Plotter、東洋測器製)でモニターすることにより、液体培地中の増殖曲線を描いた。それぞれ独立した試行を3回行った。図21に増殖曲線を示す。図21(A)は30℃で培養した結果であり、図21(B)は20℃で培養した結果である。図中、三角がCpFAH12株の増殖曲線[YEp352−CpFAH12−1、YEp352−CpFAH12−2、YEp352−CpFAH12−3]を、円がコントロール株の増殖曲線[YEp352GAP−1、YEp352GAP−2、YEp352GAP−3]を、それぞれ示す。いずれの培養温度においても、コントロール株よりもCpFAH12株のほうが、明らかに増殖が抑制されていた。すなわち、CpFAH12株の増殖のダブリングタイムは、コントロール株よりも1.4倍長くなっていた。
CpFAH12遺伝子を染色体中に組み込んだS.セレビシエの形質転換体は、CpFAH12遺伝子を染色体中に組み込んだS.ポンベの形質転換体と同様、CpFAH12を発現させることによって増殖が抑制されていた。つまり、CpFAH12遺伝子をS.セレビシエに導入することによって、S.ポンベに導入した場合と同様、リシノール酸を産生できることが示唆された。
【0061】
[実施例10]
CpFAH12遺伝子を染色体外遺伝子として導入したS.セレビシエの形質転換体を培養し、リシノール酸の産生能を調べた。対照として、S.セレビシエにYEp352GAPを導入した形質転換体を用いた。
まず、各株をNSD−URA中、20℃または30℃で一日間前培養した。次いで、培養した細胞を、前培養と同種の新しい培地で、OD600が0.05になるように希釈した後、20℃または30℃で5日間、40rpmで振とう培養した。参考例1と同様にして、培養終了後の培養液1mL当たりの各種脂肪酸の含有量を測定した。脂肪酸組成(培養液1mL当たりの各種脂肪酸の総脂肪酸含有量に対する含有割合)の結果を図22に示す。図22中、「CpFAH12」および「empty」はそれぞれ、CpFAH12を染色体外遺伝子として導入した形質転換体、およびS.セレビシエにYEp352GAPを染色体外遺伝子として導入した形質転換体を本培養した結果を示す。
図22に示すように、CpFAH12遺伝子を導入したS.セレビシエを20℃または30℃で培養した場合に、リシノール酸が産生されることがわかった。
【産業上の利用可能性】
【0062】
本発明のリシノール酸産生酵母の培養方法およびリシノール酸の製造方法により、リシノール酸を、酵母の形質転換体を用いて効率よく生産できるため、特に、リシノール酸またはひまし油が原料として使用されている化成品、塗料・印刷インキ、化粧品、医薬品、潤滑油等の製造分野において好適に用いられる。
【特許請求の範囲】
【請求項1】
FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、
前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、
を有することを特徴とするリシノール酸産生酵母の培養方法。
【請求項2】
前記低温培養工程における前記形質転換体の培養温度が、10〜33℃である請求項1に記載のリシノール酸産生酵母の培養方法。
【請求項3】
前記酵母がシゾサッカロミセス(Schizosaccharomyces)属酵母である請求項1または2に記載のリシノール酸産生酵母の培養方法。
【請求項4】
前記FAH12遺伝子が、トウゴマ(Ricinus communis)、レスクェレラ・フェンドレリ(Lesquerella fendleri)または麦角菌(Claviceps purpurea)由来である請求項1〜3のいずれか一項に記載のリシノール酸産生酵母の培養方法。
【請求項5】
請求項1〜4のいずれか一項に記載のリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法。
【請求項1】
FAH12遺伝子が導入された酵母の形質転換体を、35℃以上40℃未満で、培養液のOD600が0.5〜5.5になるまで培養する高温培養工程と、
前記高温培養工程後、前記形質転換体を35℃未満で培養する低温培養工程と、
を有することを特徴とするリシノール酸産生酵母の培養方法。
【請求項2】
前記低温培養工程における前記形質転換体の培養温度が、10〜33℃である請求項1に記載のリシノール酸産生酵母の培養方法。
【請求項3】
前記酵母がシゾサッカロミセス(Schizosaccharomyces)属酵母である請求項1または2に記載のリシノール酸産生酵母の培養方法。
【請求項4】
前記FAH12遺伝子が、トウゴマ(Ricinus communis)、レスクェレラ・フェンドレリ(Lesquerella fendleri)または麦角菌(Claviceps purpurea)由来である請求項1〜3のいずれか一項に記載のリシノール酸産生酵母の培養方法。
【請求項5】
請求項1〜4のいずれか一項に記載のリシノール酸産生酵母の培養方法により培養されたFAH12遺伝子が導入された酵母の形質転換体から、リシノール酸を取得することを特徴とするリシノール酸の製造方法。
【図4】

【図5】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図20】
【図21】
【図22】
【図1】
【図2】
【図3】
【図6】
【図19】
【図5】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図20】
【図21】
【図22】
【図1】
【図2】
【図3】
【図6】
【図19】
【公開番号】特開2013−106551(P2013−106551A)
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2011−253226(P2011−253226)
【出願日】平成23年11月18日(2011.11.18)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人科学技術振興機構、研究成果最適展開支援事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000000044)旭硝子株式会社 (2,665)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願日】平成23年11月18日(2011.11.18)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度独立行政法人科学技術振興機構、研究成果最適展開支援事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000000044)旭硝子株式会社 (2,665)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]