
形質転換植物
本発明は、葉の中にアミノ酸であるトレオニンを蓄積させる形質転換植物、具体的には、タバコ植物を提供する。トレオニンの産生を引き起こす生合成経路は、厳密に調節されており、トレオニンを過剰産出する形質転換植物を得るという以前の試みは、植物の適応度を損なってしまった。本発明は、この欠点を克服し、かつ、植物の適応度を損なうことなく、対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、植物代謝を変化させて葉の老化開始後にトレオニンの産出の増加を達成することを含む方法を見出した。本発明は、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物を含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、形質転換植物の産生に使用される遺伝子構築物に関する。この構築物は、特に葉の老化時に、植物に対してそれらの葉の中にトレオニンを蓄積させる能力を持つことができる。本発明は、そのような構築物で形質転換された植物細胞、および形質転換植物そのものに及ぶ。本発明は、形質転換植物の産生方法、および老化した植物中のトレオニン濃度を上昇させる方法にも関する。また、本発明は、遺伝子構築物で形質転換されている収穫された植物の葉(例えば、タバコの葉)、およびそのような収穫された植物の葉を含む喫煙物品にも関する。
【背景技術】
【0002】
火力乾燥されたタバコの風味を高める際の第一の目標は、アミノ酸であるトレオニンの産生である。突然変異型タバコ植物の葉の中に高濃度のトレオニンが蓄積されると、風味および香気上の大きな利点が得られる。しかし、通常は、トレオニンの産生は、アスパラギン酸族の他のアミノ酸、すなわちメチオニン、リジンおよびイソロイシンの産生と共に厳密に調節されている。従って、風味および香気上の利点を得るためには、トレオニンを産生する生合成経路の修飾が必要である。
【0003】
図1および図2に示すように、アスパラギン酸キナーゼ(AK)は、アスパラギン酸をトレオニンなどのアミノ酸に変換する植物生合成経路における最初の酵素である。内在性アスパラギン酸キナーゼは、リジンおよびトレオニンの両方からのフィードバック阻害によって調節されている。従って、フィードバック阻害を受けないアスパラギン酸キナーゼが必要である。しかし、これは、植物を無理にメチオニン飢餓状態したり、アスパラギン酸濃度をそれに依存する他の経路が制限されるまで枯渇させたりしないで達成されなければならない。
【0004】
野生型植物と比較してトレオニンを過剰産生することができる、フィードバック非感受性アスパラギン酸キナーゼを含有する形質転換植物は、成長が不十分であり、かつそのような植物が突然変異に関してホモ接合性であった場合に最悪な適応度コスト(fitness cost)を示すことが以前から分かっている。成長不良および適応度コストは、農業従事者が気付いている植物の成長における有益でない変化である。明らかに、そのような変化はいずれも望ましくない。
【0005】
従って、本発明の発明者らは、上記フィードバック阻害ループを克服するが、理想的にはその適応度に対するいかなるコストも生じさせずに葉の中にトレオニンを蓄積する形質転換植物を提供することを目的とする。これを踏まえて、本発明者らは、トレオニン非感受性アスパラギン酸キナーゼ(AK)酵素をコードする遺伝子がプロモーターの制御下にある複数の遺伝子構築物を開発して、どのような影響があるか、もし影響があれば、この遺伝子の過剰発現が老化した葉の中のトレオニン濃度に影響を与えるか否かを判定した。
【0006】
葉の老化は、葉の細胞が細胞死の前に異なる代謝および構造の変化を受ける植物の発育の一段階である。生理学的および遺伝学的研究では、老化は高度に調節されたプロセスであることが示されている。葉の老化の進行は、葉緑体の分解によって生じるクロロフィルの喪失およびその後の黄変を見かけ上の特徴とする。このような発育段階の特徴である葉のクロロフィル濃度の減少は、例えば、溶媒抽出および分光光度測定によって、あるいは、クロロフィル成分計によって測定することができる。好ましくは一定の条件下で育てられた同じ植物について記録された初期の葉のクロロフィル濃度と比較した葉のクロロフィル濃度の減少は、老化を示す。
【0007】
分子的研究では、老化は遺伝子発現における変化と関連していることが示されている。光合成に関与するタンパク質をコードするmRNAのレベルは老化時に減少するが、老化に関与していると思われているタンパク質をコードする遺伝子のmRNAレベルは増加する。老化は、老化関連遺伝子(SAG)として知られている遺伝子によって調節される高度に組織化されたプロセスである。葉の老化は、タンパク質、核酸および膜の劣化、ならびに、この劣化によってその後に起こる発育中の種子、葉または貯蔵器官などの植物の他の領域への栄養分の輸送と関連する。植物の老化の問題の1つは、老化した葉に存在する多くの有用なミネラルおよび栄養分が葉の中に残留し、そのため、葉が枯れるにつれて実質的に失われることである。老化した葉に存在する多くの他のアミノ酸と同様に、トレオニンは、枯れている葉から取り出さなければ無駄になる。
【0008】
実施例2に記載されているように、本発明者らは、植物中の特定の位置にあるトレオニン非感受性AKを発現する遺伝子構築物を用いる実験、すなわち葉特異的エンドウマメ・プラストシアニンプロモーターを用いる実験を行なった。しかし、発明者らは、そのような形質転換植物の葉は、色が淡く、肥厚し、脆くかつ帯状であることを見出した。節間部は短く、成熟度が増すにつれて褐変を示し、その芽は発育不良または奇形のいずれかであった。従って、そのような形質転換植物は、適応度コストを克服することはできない。
【0009】
そこで、本発明者らは、(SAG12プロモーターの制御下で)植物が老化に差し掛かり、よって正常に発育することが許されている後にのみ、フィードバック非感受性アスパラギン酸キナーゼ(AK)活性が発現される一連の遺伝子構築物を開発した。本発明者らは、フィードバック非感受性アスパラギン酸キナーゼ(AK)が活性化される前に、これらの構築物で形質転換された形質転換植物を成熟(すなわち、老化)まで正常に成長させることができる発明者らが開発した構築物は、意外にも適応度コストを克服することができることに気づいた。
【発明の概要】
【0010】
従って、本発明の第1の態様によれば、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物が提供される。
【0011】
本発明者らは、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に連結された老化特異的プロモーターを使用して第1の態様の構築物を形成し、次いで、これを使用して植物を形質転換させた。本発明者らは研究の結果、意外にも、老化した葉の中のトレオニン濃度が本発明に係る構築物によって上昇することを見出した。さらに、老化特異的プロモーターによって制御される導入遺伝子の発現を一時的に制限することで、トレオニン蓄積植物を産生する初期の試みで以前に認められた適応度コストという負の効果が克服される。実施例に示されているように、得られた形質転換植物は、葉の老化時に野生型の濃度よりも高いトレオニンを産生する。葉の中でトレオニンの蓄積が生じることが実証され、この濃度の上昇が当該構築物を含有するタバコの葉の風味に積極的に寄与すると考えられることから、喫煙物品をそのような葉から製造した。
【0012】
第1の態様の遺伝子構築物におけるプロモーターは、RNAポリメラーゼを誘導して、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に結合させ、かつその転写を開始できるものであればよい。
【0013】
「老化特異的プロモーター(SAG)」は、老化関連遺伝子の発現の制御に関連する任意のプロモーターとすることができる。従って、このプロモーターは、実質的に老化している組織の中でのみ、プロモーターが作動可能に連結されたコード配列(すなわち、遺伝子)の発現を制限することができる。従って、老化特異的プロモーターは、実質的に植物組織が老化している場合にのみ3’タンパク質コード領域の発現が生じるように発生学的に調節されるように、植物組織において遺伝子発現を優先的に促進することができるプロモーターとすることができる。当然のことながら、老化は古い葉などの植物の古い部分で生じ、種子などの植物の若い部分では生じない傾向がある。
【0014】
多数の老化関連遺伝子を発現することで知られている植物の一例は、シロイヌナズナ(Arabidopsis)である。従って、第1の態様に係る構築物に存在するプロモーターは、シロイヌナズナの老化関連遺伝子から単離したものであってもよい。Gepsteinら(The Plant Journal, 2003, 36, 629-642)は、モデルとしてシロイヌナズナを用いて、SAGおよびそのプロモーターについて詳しい研究を行なった。従って、当該遺伝子構築物は、本明細書に開示されている任意のSAG由来のプロモーターを含んでいてもよい。例えば、好適なプロモーターは、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは機能的な断片から選択されてもよい。
【0015】
好ましいプロモーターは、SAG12およびSAG13プロモーターである。一実施形態では、当該プロモーターは、当業者に公知であるSAG12プロモーターまたはその機能的な変異体もしくは断片である(Gan & Amasino, 1997, Plant Physiology, 113: 313-319)。SAG12プロモーターをコードするDNA配列が図6に示されており、本明細書では以下のとおり配列番号1と称する:
配列番号1
TCGAGACCCGATTGTTATTTTTAGACTGAGACAAAAAAGTAGAATCGTTGATTGTTAAAATTTAAAATTAGTTTCATTACGTTTCGATAAAAAAATGATTAGTTTATCATAGCTTAATTATAGCATTGATTTCTAAATTTGTTTTTTGACCACCCTTTTTTCTCTCTTTGGTGTTTTCTTAACATTAGAAGAACCCATAACAATGTACGTTCAAATTAATTAAAAACAATATTTCCAAGTTTTATATACGAAACTTGTTTTTTTTAATGAAAACAGTTGAATAGTTGATTATGAATTAGTTAGATCAATACTCAATATATGATCAATGATGTATATATATGAACTCAGTTGTTATACAAGAAATGAAAATGCTATTTAAATACAGATCATGAAGTGTTAAAAAGTGTCAGAATATGACATGAAGCGTTTTGTCCTACCGGGTATTCGAGTTATAGGTTTGGATCTCTCAAGAATATTTTGGGCCATACTAGTTATATTTGGGCTTAAGCGTTTTGCAAAGAGACGAGGAAGAAAGATTGGGTCAAGTTAACAAAACAGAGACACTCGTATTAGTTGGTACTTTGGTAGCAAGTCGATTTATTTGCCAGTAAAAACTTGGTACACAACTGACAACTCGTATCGTTATTAGTTTGTACTTGGTACCTTTGGTTCAAGAAAAAGTTGATATAGTTAAATCAGTTGTGTTCATGAGGTGATTGTGATTTAATTTGTTGACTAGGGCGATTCCTTCACATCACAATAACAAAGTTTTATAGATTTTTTTTTTATAACATTTTTGCCACGCTTCGTAAAGTTTGGTATTTACACCGCATTTTTCCCTGTACAAGAATTCATATATTATTTATTTATATACTCCAGTTGACAATTATAAGTTTATAACGTTTTTACAATTATTTAAATACCATGTGAAGATCCAAGAATATGTCTTACTTCTTCTTTGTGTAAGAAAACTAACTATATCACTATAATAAAATAATTCTAATCATTATATTTGTAAATATGCAGTTATTTGTCAATTTTGAATTTAGTATTTTAGACGTTATCACTTCAGCCAAATATGATTTGGATTTAAGTCCAAAATGCAATTTCGTACGTATCCCTCTTGTCGTCTAATGATTATTTCAATATTTCTTATATTATCCCTAACTACAGAGCTACATTTATATTGTATTCTAATGACAGGGAAACCTTCATAGAGATTCAGATAGATGAAATTGGTGGGAAACATCATTGAACAGGAAACTTTTAGCAAATCATATCGATTTATCTACAAAAGAATACGTAGCGTAATGAAGTCCACTTGTTGTGAATGACTATGATTTGATCAAATTAGTTAATTTTGTCGAATCATTTTTCTTTTTGATTTGATTAAGCTTTTAACTTGCACGAATGGTTCTCTTGTGAATAAACAGAATCTTTGAATTCAAACTATTTGATTAGTGAAAAGACAAAAGAAGATTCCTTGTTTTTATGTGATTAGTGATTTTGATGCATGAAAGGTACCTACGTACTACAAGAAAAATAAACATGTACGTAACTACGTATCAGCATGTAAAAGTATTTTTTTCCAAATAATTTATACTCATGATAGATTTTTTTTTTTTGAAATGTCAATTAAAAATGCTTTCTTAAATATTAATTTTAATTAATTAAATAAGGAAATATATTTATGCAAAACATCATCAACACATATCCAACTTCGAAAATCTCTATAGTACACAAGTAGAGAAATTAAATTTTACTAGATACAAACTTCCTAATCATCAAATATAAATGTTTACAAAACTAATTAAACCCACCACTAAAATTAACTAAAAATCCGAGCAAAGTGAGTGAACAAGACTTGATTTCAGGTTGATGTAGGACTAAAATGACTACGTATCAAACATCAACGATCATTTAGTTATGTATGAATGAATGTAGTCATTACTTGTAAAACAAAAATGCTTTGATTTGGATCAATCACTTCATGTGAACATTAGCAATTACATCAACCTTATTTTCACTATAAAACCCCATCTCAGTACCCTTCTGAAGTAATCAAATTAAGAGCAAAAGTCATTTAACTTAGG
従って、本発明の構築物におけるプロモーターは、実質的に配列番号1に示されているようなヌクレオチド配列またはその機能的な変異体もしくは機能的な断片を含んでいてもよい。SAG12プロモーター配列は、米国特許第5,689,042号に記載されているようなシロイヌナズナ(Arabidopsis thaliana)から得られたものであってもよい。プロモーターがSAG12である実施形態では、当然のことながら、プロモーターは、配列番号1の1〜2093の各塩基を含んでいてもよい。ただし、プロモーターの機能的な変異体もしくは機能的な断片が本発明の遺伝子構築物に使用されてもよい。
【0016】
「プロモーターの機能的な変異体もしくは機能的な断片」は、そこに作動可能に連結された任意のコード領域の発現を開始させるのに機能的に十分である誘導体またはプロモーターの一部とすることができる。例えば、プロモーターがSAG12に基づいている実施形態では、当業者であれば、配列番号1が修飾されても、SAG12プロモーターの部分のみが必要とされてもよく、従って、当該構築物において、それらがトレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドの遺伝子発現をなお開始させるということを理解しているであろう。同様の修飾を、SAG13、SAG101、SAG21およびSAG18などの他の公知のSAGプロモーターのいずれかのヌクレオチド配列に対して行なってもよい。
【0017】
プロモーターの機能的な変異体および機能的な断片は、転写酵素が推定プロモーター領域に結合し、次いで、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドのコード配列の転写を引き起こすか否かを評価することによって容易に同定することができる。あるいは、そのような機能的な変異体および断片は、コード領域に関連づけられている場合にはプロモーター上で突然変異生成を行ない、遺伝子発現が生じ得るか否かを評価することによって調べてもよい。
【0018】
第1の態様の遺伝子構築物は、老化時にトレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドの発現を引き起こし得るものであればよい。このプロモーターは、トレオニン非感受性アスパラギン酸キナーゼ活性を示すポリペプチドをコードするコード配列の発現を誘導してもよい。従って、当該遺伝子構築物は、トレオニン非感受性アスパラギン酸キナーゼ(AK)をコードする少なくとも1つのコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。従って、第1の実施形態では、遺伝子構築物は、老化特異的プロモーター、およびトレオニン非感受性アスパラギン酸キナーゼ(AK)をコードするコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0019】
実施例3〜4に記載されているように、本発明者らは、本発明の構築物によって植物を形質転換することによって宿主細胞においてトレオニン非感受性アスパラギン酸キナーゼを発現させると、トレオニン濃度の著しい上昇が引き起こされることを見出した。さらに、有利なことに、本発明者らは、本発明の構築物が、形質転換された植物の適応度に対する有害作用を全く有しないことを見出した。
【0020】
図1および図2に示すように、植物では、アミノ酸であるリジン、トレオニン、メチオニンおよびイソロイシンは、アスパラギン酸から合成される。いくつかのフィードバック阻害ループがこの経路で認められている。この経路での最初の酵素であるアスパラギン酸キナーゼ(AK)(EC 2.7.2.4)は、アスパラギン酸のリン酸化を触媒して、付随して生じるATPの加水分解によって3−アスパルチルリン酸を生成する。高等な植物は一般に、少なくとも2種または3種のAKイソ酵素を有すると考えられている。AK活性は、最終産物であるアミノ酸(リジンおよびトレオニン)から負のフィードバックを受ける。少なくとも1種のAKイソ酵素はトレオニンによって、もう1つはリジンおよびS−アデノシルメチオニンによってフィードバック阻害される。オオムギでは、AK活性は、3つのイソ酵素ピーク(isozymal peak)に分類することができ、その内の1つはトレオニンによって阻害され、他の2つはリジンによって阻害される。さらなるフィードバックループは、生合成経路において、ジヒドロピコリン酸合成酵素(DHPS)およびホモセリン不飽和化酵素(HSD)などの他の酵素により作動する。
【0021】
図1および図2に示すように、ホモセリン脱水素酵素(HSDH)としても知られているホモセリン不飽和化酵素(HSD)(EC 1.1.1.3)は、トレオニン、メチオニンおよびイソロイシンの生合成、すなわち3−アスパラギン酸セミアルデヒドのホモセリンへの変換に一意的に関連づけられている第1の反応を触媒する。高等な植物は一般に、少なくとも2つの型のHSDH、すなわちトレオニン感受性型およびトレオニン非感受性型を有する。精製HSDHおよびcDNAクローンの特性評価では、HSDHイソ酵素は単一のタンパク質上のAKに結びづけられていることが確認されており、よって「AK:HSDH」と表わされる。
【0022】
葉のトレオニン(Thr)含有量の上昇は、合成経路への負のフィードバック制御を克服するか否かによって決まる。例えば、大腸菌由来のリジン非感受性AKを発現している形質転換タバコが産生されており、これはトレオニン蓄積表現型を示した。この形質転換体では、細胞質または葉緑体のいずれかを標的にした細菌性AKがタバコにおいて35Sの制御下で発現された。それでも、AKの内因性活性は、リジンおよびトレオニンの両方による阻害をまったく受けなかった。葉緑体を標的とした導入遺伝子は、より高いAK活性を示した。葉緑体型を発現している植物ではより高いトレオニン濃度が認められた。しかし、植物の成長不良が顕著であり、ホモ接合植物は、しわの寄った上葉、開花の遅延および部分的な不稔を含む適応度コストを示した。
【0023】
シロイヌナズナにおいて行われた研究(Paris et al, 2003, The Journal of Biological Chemistry, Vol 278, no. 7, pp5361-5366)では、AK:HSDH酵素の調節領域が、一般的なループ−αへリックス−ループ−βスランド−ループ−βスランドモチーフによって定義される2つの相同サブドメインを含むことが実証されている。部位特異的突然変異誘発が、トレオニン結合部位を解明するために使用された。アスパラギン酸キナーゼ−ホモセリン脱水素酵素のモノマーの各調節領域は、部分的にGln443およびGln524によって構成された2つの非等価トレオニン結合部位を有することが見出された。トレオニンのGln443への結合はAK活性を阻害し、さらに2つ目のトレオニンのGln524への結合を容易にして、それにより、HSDH阻害を引き起こす。
【0024】
図3は、トレオニンによるAK−HSDHの阻害のための提案モデルを示す。ここでは、AKおよびHSDHの活性触媒領域は四角で表わされており、阻害された触媒領域は三角で表わされている。1つ目のサブドメインへのトレオニン(Thr)結合は、(i)他のサブドメインの立体構造変化、および(ii)AK触媒領域の立体構造変化の両方を導き、それによりAK阻害が引き起こされる。2つ目のサブドメインの立体構造変化は、2つ目のトレオニンの結合を誘導し、それによりHSDH触媒領域の立体構造変化およびHSDH阻害が引き起こされる。
【0025】
これらのグルタミン残基のアラニンへの突然変異によって酵素のトレオニン阻害を無効にさせる。この突然変異は、HSDH活性の動態には影響を与えず、トレオニンに対するその感受性のみに影響を与える。すなわち、AK動態は僅かに変化するだけである。しかし、残念なことに、シロイヌナズナ由来のフィードバック非感受性AK:HSDHが導入されかつ発現される形質転換植物は、適応度コストも引き起こす。
【0026】
対照的に、本発明の遺伝子構築物は、老化時にトレオニン非感受性アスパラギン酸活性を有するポリペプチドの発現を引き起こし、形質転換植物の適応度に対する有害作用を全く示さなかった。従って、第1の態様の遺伝子構築物は、トレオニン非感受性アスパラギン酸キナーゼ(AK)、またはトレオニン非感受性の二機能性アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)、あるいはその機能的な変異体もしくは機能的な断片をコードしてもよい。
【0027】
トレオニン非感受性AKまたは二機能性AK−HSDH、あるいはその機能的な変異体もしくは断片は、植物などの任意の好適な供給源に由来するものであってもよい。トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ属、トウモロコシ(Zea)属、フラベリア(Flaveria)属またはクレオメ(Cleome)属に由来するものであってもよい。トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ(Arabidopsis thaliana)、トウモロコシ(Zea mays)、フラベリア・トリネルビア(Flaveria trinervia)、フラベリア・ビデンティス(Flaveria bidentis)、フラベリア・ブラウニー(Flaveria brownie)またはフウチョウソウ(Cleome gynandra)に由来するものであってもよい。好ましくは、当該酵素のコード配列は、シロイヌナズナ(Arabidopsis thaliana)などのシロイヌナズナ(Arabidopsis)属に由来するものであってもよい。
【0028】
特に好ましいトレオニン非感受性酵素は、少なくとも1つのトレオニン結合部位が変化している突然変異型AK−HSDHである。好ましくは、部分的にGln443およびGln524で構成されているトレオニン結合部位の一方または両方が突然変異されている。例えば、シロイヌナズナのAK−HSDHは、Gln443および/またはGln524で突然変異されていてもよい。好ましくは、シロイヌナズナのAK−HSDHは、Gln443およびGln524で突然変異されている。本発明で使用される突然変異型AK:HSDH遺伝子が図7に示されており、ここでは、突然変異型塩基に下線が引かれている。
【0029】
従って、シロイヌナズナのトレオニン非感受性アスパラギン酸キナーゼの一実施形態(すなわち、Gln443Ala単一突然変異体)をコードするDNA配列が、以下のとおり配列番号2として以下に規定される:
配列番号2
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTCARGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号2(すなわち、Q443A)では、強調されているGCTはアラニンをコードする突然変異型Gln443に対応し、強調されているCARは野生型Gln524に対応する。ここで、RはGまたはAのいずれであってもよい。
【0030】
シロイヌナズナトレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln524Ala単一突然変異体)をコードするDNA配列が、以下のとおり配列番号3として以下に規定される:
配列番号3
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCACARGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号3(すなわち、Q524A)では、強調されているGCTはアラニンをコードする突然変異型Gln524に対応し、強調されているCARは野生型Gln443に対応する。ここで、RはGまたはAのいずれであってもよい。
【0031】
シロイヌナズナトレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln443Ala;Gln524Ala二重突然変異体)をコードするDNA配列は、以下のとおり配列番号4として以下に規定される:
配列番号4
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号4(すなわち、Q443A;Q524A)では、強調されているGCTは、どちらもアラニンをコードする突然変異型Gln443およびGln524に対応する。
【0032】
従って、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、実質的に配列番号2、3または4のうちのいずれか1つに示されているような核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0033】
トレオニン非感受性アスパラギン酸キナーゼの一実施形態(すなわち、Gln443Ala単一突然変異体)のポリペプチド配列が、以下のとおり配列番号5として以下に規定される:
配列番号5
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISAASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISQGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGG
IFSDILRLASYLGAPS 配列番号5では、位置443にある突然変異型アラニン(A)、および位置524にある野生型グルタミン(Q)が強調されている。
【0034】
トレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln524Ala単一突然変異体)のポリペプチド配列は、以下のとおり配列番号6として以下に規定される:
配列番号6
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISQASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISAGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGGIFSDILRLASYLGAPS
配列番号6では、位置524にある突然変異型アラニン(A)および位置443にある野生型グルタミン(Q)が強調されている。
【0035】
トレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln443Ala;Gln524Ala二重突然変異体)のポリペプチド配列は、以下のとおり配列番号7として以下に規定される:
配列番号7
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISAASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISAGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGGIFSDILRLASYLGAPS
配列番号7では、位置443および524にある突然変異型アラニン(A)が強調されている。
【0036】
従って、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチド配列は、実質的に配列番号5、6または7のうちのいずれか1つに示されているようなアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0037】
本発明の遺伝子構築物は、発現カセットの形態であってもよく、宿主細胞におけるコード配列の発現に適したものであればよい。本発明の遺伝子構築物は、それがベクターに組み込まれることなく、宿主細胞に導入されてもよい。例えば、核酸分子であり得る遺伝子構築物は、リポソームまたはウイルス粒子内に組み込まれてもよい。あるいは、精製された核酸分子(例えば、ヒストンを含まないDNA、すなわち裸のDNA)が、例えば、直接的なエンドサイトーシス取り込みなどの好適な手段によって宿主細胞に直接挿入されてもよい。当該遺伝子構築物は、形質移入、感染、微量注入、細胞融合、原形質融合または衝撃照射(ballistic bombardment)によって宿主対象(例えば、植物)の細胞に直接導入されてもよい。あるいは、本発明の遺伝子構築物は、パーティクルガン法を用いて宿主細胞に直接導入されてもよい。あるいは、当該遺伝子構築物は、好適な宿主細胞における発現のために組換え型ベクター内部に含まれていてもよい。
【0038】
従って、第2の態様では、第1の態様に係る遺伝子構築物を含む組換え型ベクターが提供される。
【0039】
当該組換え型ベクターは、プラスミド、コスミドまたはファージであってもよい。そのような組換え型ベクターは、宿主細胞を第1の態様の遺伝子構築物で形質転換し、かつその中の発現カセットを複製するのに非常に有用である。当業者であれば、本発明の遺伝子構築物は、発現のために、多くの種類のバックボーンベクターと組み合わせ得ることを理解しているであろう。バックボーンベクターは、例えば、1つのベクターが大腸菌およびアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)の両方で複製することができるバイナリーベクターであってもよい。例えば、好適なベクターは、pBIN19などのpBINプラスミドであってもよい。しかし、好ましいバックボーンベクターはBNP1380000001であり、これは、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290) に基づいており、かつSAG12プロモーターを内部に含む。このベクターの一実施形態が図16に示されている。
【0040】
組換え型ベクターは、プロモーター(例えば、老化関連プロモーター)に加えて、様々な他の機能的な成分、および少なくとも1つの(突然変異型AK−HSDHをコードする)コード配列を含んでいてもよい。例えば、組換え型ベクターは、宿主細胞中の細胞質ゾル内で自己複製するように設計されていてもよい。この場合、組換え型ベクター内に、DNA複製を誘導または調節する成分が必要な場合がある。あるいは、組換え型ベクターは、宿主細胞のゲノムに組み込まれるように設計されていてもよい。この場合、(例えば相同組換えによる)標的組換えを支持するDNA配列が想定される。
【0041】
また、当該組換え型ベクターは、クローニングプロセスにおける選択可能なマーカーとして、すなわち、形質移入または形質転換されている細胞の選択を可能にするため、および異種のDNAが組み込まれているベクターを内部に含む細胞の選択を可能にするために使用できる遺伝子のDNAコードを含んでいてもよい。あるいは、選択可能なマーカー遺伝子は、目的の遺伝子を含有するベクターと同時に使用される異なるベクター中に存在していてもよい。当該ベクターは、コード配列の発現の調節に関与するDNA、または発現されるポリペプチドをある種の宿主細胞、例えば葉緑体に導くためのDNAを含んでいてもよい。従って、第2の態様のベクターは、選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子)、ポリペプチド終結シグナルおよびタンパク質標的配列(例えば、葉緑体輸送ペプチド)からなる群から選択される少なくとも1つのさらなる成分を含んでいてもよい。
【0042】
好適なマーカー遺伝子の例としては、抗生物質耐性遺伝子、例えば、カナマイシン、ジェネテシン(G418)およびハイグロマイシン(npt−II、hyg−B)に対して耐性を示す遺伝子;除草剤耐性遺伝子、例えば、ホスフィノトリシンおよびスルホンアミド系除草剤(それぞれbarおよびsuI;欧州特許出願公開第242246号、欧州特許出願公開第0249637号)に対して耐性を示す遺伝子;およびスクリーニング可能なマーカー、例えば、β−グルクロニダーゼ(英国特許出願公開第2197653号)、ルシフェラーゼおよび緑色蛍光タンパク質(GFP)が挙げられる。
【0043】
当該マーカー遺伝子は、第2のプロモーター(老化関連プロモーターであってもそうでなくてもよい)によって制御されてもよく、細胞における発現を可能にし、種子の中に存在するものであってもそうでなくてもよく、それにより、植物の発育の任意の段階でマーカーを含有する細胞または組織の選択を可能にする。好適な第2のプロモーターは、アグロバクテリウム(Agrobacterium)のノパリン合成酵素遺伝子のプロモーターおよび35Sカリフラワーモザイクウイルス(CaMV)転写物をコードする遺伝子に由来するプロモーターである。ただし、任意の他の好適な第2のプロモーターが使用されてもよい。
【0044】
本発明の遺伝子構築物の様々な実施形態は、実施例2に記載されていて、かつ以下のように要約することができる好適なクローニング法を用いて産生されてもよい。野生型AK−HSDHをコードする遺伝子は、好適なプライマーを用いるPCRによってゲノムまたはcDNAの鋳型のいずれかから増幅されてもよい。野生型AK−HSDH遺伝子の増幅に適したプライマーは、配列番号8および/または配列番号9であってもよい。PCR産物は、アガロースゲル電気泳動法を用いて調べてもよい。次いで、位置443および/または524にある野生型コドンを突然変異させてGln443AlaおよびGln524Ala単一突然変異体または二重突然変異体を産生するために、好適な対のプライマーを用いる部位特異的突然変異誘発を行なってもよい。例えば、Gln443のコドンを変化させるのに適したプライマーは、配列番号10および/または配列番号11であってもよい。Gln524のコドンを変化させるのに適したプライマーは、配列番号12および/または配列番号13であってもよい。
【0045】
次いで、この2つの単一突然変異体または二重突然変異体のうちのいずれか一方をコードするPCR産物を、クローニングに適したベクター、例えば、Invitrogen社から入手でき、pCR4 Blunt-TOPOベクターという商品名で市販されているベクターの中に連結してもよい。次いで、PCR産物を内部に含むベクターを大腸菌などの好適な宿主中で成長させてもよい。次いで、大腸菌コロニーを、好適なプライマーを用いるPCRによってスクリーニングしてもよく、正しい制限酵素消化パターンを示すプラスミド中の挿入断片を好適なプライマーを用いて配列決定してもよい。
【0046】
TOPO−cDNA(AK−HSDH)を保持する大腸菌コロニーを培養して好適な量のプラスミドを作製し、次いで、これを精製してもよい。ついで、プラスミドを温浸して、突然変異型AK−HSDHをコードするDNA断片を遊離させた後、これを、pBNPプラスミドなどの、好適なプロモーター(例えば、SAGプロモーター(好ましくは、SAG12))を内部に含むベクターにクローン化してもよい。得られたプラスミドは、pBNP138−0453−001と命名された。第2の態様に係るベクターの一実施形態は、実質的に図15に示されているようなものであってもよい。
【0047】
第3の態様では、同じ条件下で栽培された対応する野生型植物よりも高濃度のトレオニンを葉に蓄積させる形質転換植物の産生方法が提供される。当該方法は、
(i)植物細胞を、第1の態様の遺伝子構築物または第2の態様のベクターで形質転換することと、
(ii)形質転換された細胞から植物を再生することと、
を含む。
【0048】
植物の葉の中のトレオニン濃度および植物の成長率の測定方法が、実施例1に記載されている。第3の態様の方法は、試験植物細胞を第1の態様に係る遺伝子構築物または第2の態様に係るベクターで形質転換することを含む。当該遺伝子構築物またはベクターは、任意の好適な手段によって宿主細胞に導入してもよい。
【0049】
第4の態様では、第1の態様に係る遺伝子構築物または第2の態様に係る組換え型ベクターを含む細胞が提供される。
【0050】
当該細胞は植物細胞であってもよい。本発明者らは、宿主細胞における老化特異的プロモーターの制御下でのトレオニン非感受性アスパラギン酸キナーゼの発現は、意外にも、適応度を損なうことなく老化した葉の中のトレオニン濃度を上昇させるのに有効であることに気づいたため、第4の態様の細胞は、1つまたは複数の第1の態様の構築物あるいは1つまたは複数の第2の態様のベクターを含んでいてもよい。
【0051】
当該細胞は、公知の技術を用いて本発明に係る遺伝子構築物またはベクターで形質転換されていてもよい。遺伝子構築物を宿主細胞に導入するのに適した手段は、例えば、欧州特許出願公開第0116718号および第0270822号に記載されているように当該技術分野で知られている手段による、アグロバクテリウムが保持するディスアーム型Tiプラスミドベクターの使用を含んでもよい。さらなる方法は、植物原形質を形質転換することであってもよく、この方法では、最初に細胞壁を除去し、核酸を導入し、次いで、細胞壁を再編成する。次いで、形質転換された細胞を植物に成長させてもよい。
【0052】
第5の態様では、第1の態様に係る遺伝子構築物または第2の態様に係るベクターを含む形質転換植物が提供される。
【0053】
第5の態様に係る形質転換植物としては、アブラナ属などのアブラナ科が挙げられる。当該植物は、セイヨウアブラナ(Brassica napus)(菜種)であってもよい。
【0054】
第5の態様に係る形質転換植物のさらなる例としては、コムギ連(Triticeae)などのイネ科が挙げられる。当該植物は、コムギ属(Triticum)(コムギ)であってもよい。コムギ中の種子タンパク質含有量を増加させることによって、パンなどのそのようなコムギを含む食品の体積を増加させてもよい。
【0055】
第5の態様に係る好適な形質転換植物のさらなる例としては、植物のナス科が挙げられ、ナス科の例としては、例えば、チョウセンアサガオ、ナス、マンドレイク、ベラドンナ(deadly nightshade (belladonna))、トウガラシ(パプリカ、カラトウガラシ)、ジャガイモおよびタバコが挙げられる。ナス科の好適な属の一例はニコチアナである。ニコチアナの好適な種は、タバコ植物、または単にタバコと称される場合がある。植物を第1の態様の遺伝子構築物または第2の態様のベクターで形質転換する様々な方法が知られており、本発明で使用することができる。
【0056】
例えば、タバコは、以下のように形質転換してもよい。タバコ(Nicotiana tabacum)を、基本的にHorsch et al. (Science 227: 1229-1231, 1985)に記載されているように葉片共培養法を用いて形質転換した。最も若い2枚の広がった葉を7週齢のタバコ植物から採取してもよく、8%のドメストス(商標)中で10分間表面滅菌し、滅菌蒸留水で6回洗浄してもよい。葉片をNo.6コルク穿孔器を用いて切断し、約2分間かけて(本発明に係る)適当なバイナリーベクターを含有するアグロバクテリウム懸濁液に入れてもよい。この葉片を、2枚の無菌濾紙の間に穏やかにブロットしてもよい。10枚の葉片をLS3%スクロース+2μMのBAP+0.2μMのNAAを含むプレート上に置き、次いで、これを成長室(growth room)において2日間インキュベートしてもよい。
【0057】
葉片は、クラフォラン500g/lおよびのカナマイシン100g/lを添加したLS+3%スクロース+2μMのBAP+0.2μMのNAAを含むプレートに移してもよい。この葉片は、2週間後に上記培地を含む新しいプレートに移してもよい。さらに2週間後、この葉片は、クラフォラン500mg/lおよびカナマイシン100mg/lを添加したLS+3%スクロース+0.5μMのBAPを含むプレートに移してもよい。この葉片は、2週間ごとに新しい培地に移してもよい。苗条が現れたら、それらを切除し、クラフォラン500mg/lを添加したLS+3%スクロースを入れた瓶に移してもよい。瓶の中の苗条は、約4週間後、LS+3%スクロース+250mg/lのクラフォランに移してもよい。さらに3〜4週間後、この植物を、LS+3%スクロース(抗生物質なし)に移して、根づかせてもい。植物が根づいたら、温室の土壌に移してもよい。
【0058】
第6の態様では、第5の態様に係る形質転換植物から得られた植物の繁殖用製品(propagation product)が提供される。
【0059】
「植物繁殖用製品」は、そこからさらなる植物が生産することができる任意の植物とすることができる。好適には、この植物繁殖用製品は種子であってもよい。
【0060】
また、本発明は、同じ条件下で栽培された対応する野生型植物から収穫された葉よりも高濃度のトレオニンを含有する、本発明の形質転換植物から収穫された葉も包含する。
【0061】
従って、第7の態様では、同じ条件下で栽培された野生型植物から採取された、収穫された葉の中の対応するトレオニン濃度よりも高濃度のトレオニンを含有する収穫された葉が提供され、ここで、この葉は、第5の態様に係る形質転換植物から収穫されたものか、第3の態様に係る方法によって生産されたものである。
【0062】
本発明の第8の態様は、老化した葉の中でトレオニンを過剰産出することができる突然変異型タバコ植物から得られたトレオニンが豊富なタバコを含む喫煙物品を提供する。
【0063】
有利かつ好ましくは、突然変異型タバコ植物は、本発明の遺伝子構築物またはベクターで形質転換されたものであってもよい。当該喫煙物品は、紙巻タバコ、葉巻タバコ、シガリロ(小型葉巻)または手巻きタバコなどであってもよい。
【0064】
トレオニンが減少したタバコには、同じ条件下で栽培された野生型植物中の対応する濃度よりもトレオニン濃度の高いタバコが含まれる。そのような喫煙物品は、突然変異型タバコ植物から得られたタバコを含んでいてもよく、このタバコは、本発明の第1の態様に係る遺伝子構築物または第2の態様に係るベクターで形質転換されたものであってもよい。トレオニンが豊富なタバコは風味および香気がよくなっている。
【0065】
当然のことながら、本発明は、植物の適応度を損なうことなく、対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、植物の代謝を変化させて、葉の老化開始後にトレオニンの産生の増加を達成することを含む方法を提供する。
【0066】
従って、本発明の第9の態様では、植物の適応度を損なうことなく、対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、葉の老化開始後にトレオニンの産生の増加を達成することを含む方法が提供される。
【0067】
好ましくかつ有利なことには、本発明に係る方法は、生産される植物の健康または適応度を損なうことはない。好ましくは、当該方法は、第1の態様の遺伝子構築物または第2の態様のベクターを用いて、試験植物および好ましくはその葉を形質転換することを含む。
【0068】
実施例4に記載されているように、本発明の形質転換された植物中のトレオニン濃度を測定し、かつ老化した葉の中のトレオニン濃度の上昇を示すことに加えて、本発明者らは、形質転換植物中のグルタミン、グルタミン酸、アスパラギン酸およびヒスチジンなどの他のアミノ酸の濃度も測定した。図10〜図14に示すように、これらの各アミノ酸の濃度は、対照試料に匹敵するかそれのよりも高く、これは、形質転換植物の適応度が損なわれていなかったことを強く示している。従って、要約すると、本発明の構築物は、形質転換植物の適応度に悪影響を与えることなく、老化した葉の中のトレオニン濃度を上昇させるために好適に使用することができる。
【0069】
当然のことながら、本発明は、実質的に本明細書で参照されている配列のいずれかのアミノ酸または核酸配列(その機能的な変異体もしくは機能的な断片を含む)を含む、任意の核酸、ペプチド、その変異体、誘導体または類似体にまで及ぶ。「実質的にアミノ酸/ポリヌクレオチド/ポリペプチド配列」、「機能的な変異体」および「機能的な断片」という用語は、本明細書で参照されている配列のうちのいずれか1つのアミノ酸/ポリヌクレオチド/ポリペプチド配列との少なくとも40%の配列同一性、例えば、配列番号1(すなわちSAG12プロモーター)として同定されているプロモーターまたは(AK−HSDH酵素の様々な実施形態をコードする)配列番号2、3または4として同定されている遺伝子との40%の同一性、あるいは配列番号5、6または7(すなわち、突然変異型AK−HSDH酵素の様々な実施形態)として同定されているポリペプチドとの40%の同一性などを有する配列とすることができる。
【0070】
また、参照されている配列のいずれかに対して65%を超える、より好ましくは70%を超える、さらにより好ましくは75%を超える、なおより好ましくは80%を超える配列の同一性を有するアミノ酸/ポリヌクレオチド/ポリペプチド配列も想定される。好ましくは、アミノ酸/ポリヌクレオチド/ポリペプチド配列は、参照されている配列のいずれかとの少なくとも85%の同一性、より好ましくは少なくとも90%の同一性、さらにより好ましくは少なくとも92%の同一性、なおより好ましくは少なくとも95%の同一性、なおより好ましくは少なくとも97%の同一性、なおより好ましくは少なくとも98%の同一性、最も好ましくは本明細書で参照されている配列のいずれかとの少なくとも99%の同一性を有する。
【0071】
当業者であれば、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算方法が分かっているであろう。2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合を計算するために、最初に2つの配列のアラインメントを作成した後、配列の同一性値を計算しなければならない。2つの配列の同一性の割合は、(i)配列を整列させるために使用される方法、例えば、ClustalW、BLAST、FASTA、Smith−Waterman法(異なるプログラムにより実行される)、あるいは3D比較による構造アラインメント、および(ii)アラインメント法(例えば、ローカルまたはグローバルアラインメント)によって使用されるパラメータ、使用される対のスコア行列(例えば、BLOSUM62、PAM250、Gonnetなど)、およびギャップペナルティ(例えば、関数および定数)によって異なる値を取る可能性がある。
【0072】
アラインメント作成後の、2つの配列間の同一性の割合に関する計算方法は数多くある。例えば、(i)最も短い配列の長さ、(ii)アラインメントの長さ、(iii)配列の平均長、(iv)ギャップのない位置の数、または(iv)オーバーハングを除く等価位置の数によって同一部分の数を割ってもよい。さらに、当然のことながら、同一性の割合は長さにも強く依存する。従って、一対の配列が短い程、偶然に生じることを予測し得る配列の同一性が高くなる。
【0073】
従って、当然のことながら、タンパク質またはDNA配列の正確なアラインメントは、複雑なプロセスである。有名なマルチプルアライメントプログラムであるClustalW(Thompson et al., 1994, Nucleic Acids Research, 22, 4673-4680; Thompson et al., 1997, Nucleic Acids Research, 24, 4876-4882)は、本発明に従ってタンパク質またはDNAのマルチプルアラインメントを生成する好ましい方法である。ClustalWの好適なパラメータは、以下のとおりであってもよい:DNAアラインメントについては:ギャップ開始ペナルティー(Gap Open Penalty)=15.0、ギャップ伸長ペナルティー(Gap Extension Penalty)=6.66および行列=同一性。タンパク質アラインメントについては:ギャップ開始ペナルティー=10.0、ギャップ伸長ペナルティー=0.2および行列=Gonnet。DNAおよびタンパク質アラインメントについては:ENDGAP=−1およびGAPDIST=4。当業者であれば、最適な配列アラインメントのためにこれらのパラメータおよび他のパラメータを変える必要があるかも知れないことが分かっているであろう。
【0074】
好ましくは、次に、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算は、(N/T)*100(式中、Nは、同一の残基を共有する配列の位置の数であり、Tは、ギャップを含めるがオーバーハングを除いて比較される位置の合計数である)としてそのようなアラインメントから計算してもよい。従って、2つの配列間の同一性の割合の最も好ましい計算方法は、(i)好適な組み合わせのパラメータ(例えば、上記のようなパラメータ)を用いるClustalWプログラムを用いて配列アラインメントを作成すること、および(ii)NおよびTの値を式:配列の同一性=(N/T)*100に書き込むことを含む。
【0075】
同様の配列の他の同定方法が当該技術分野で知られている。例えば、実質的に同様のヌクレオチド配列は、厳密な条件下で、配列番号1、2、3または4に示されている配列あるいはそれらの相補体とハイブリッドを形成する配列によってコードされるであろう。厳密な条件とは、ヌクレオチドが約45℃で3倍の塩化ナトリウム/クエン酸ナトリウム(SSC)中でフィルター結合DNAまたはRNAとハイブリッド形成した後、約20〜65℃で0.2倍のSSC/0.1%SDS中で少なくとも1回洗浄されることを意味する。あるいは、実質的に同様のポリペプチドは、配列番号5、6または7に示されている配列とは、少なくとも1個、ただし5、10、20、50または100個未満のアミノ酸が異なっていてもよい。
【0076】
遺伝子コードの縮重により、本明細書に記載されている任意の核酸配列を、それによってコードされるタンパク質の配列に実質的に影響を与えることなく変化または変更させてその機能的な変異体を得ることができることは明らかである。好適なヌクレオチド変異体は、配列内の同じアミノ酸をコードする異なるコドンの置換によって静的な変化(silent change)を生じさせることにより変化した配列を有する変異体である。他の好適な変異体は、相同ヌクレオチド配列を有するが、置換されたアミノ酸と同様の生物物理学的特性の側鎖を有するアミノ酸をコードする異なるコドンの置換によって保存的変化(conservative change)を生じさせることにより変化した配列の全てもしくは部分を含む変異体である。例えば、小型で非極性の疎水性アミノ酸としては、グリシン、アラニン、ロイシン、イソロイシン、バリン、プロリンおよびメチオニンが挙げられる。大型で非極性の疎水性アミノ酸としては、フェニルアラニン、トリプトファンおよびチロシンが挙げられる。中立極性のアミノ酸としては、セリン、トレオニン、システイン、アスパラギンおよびグルタミンが挙げられる。正電荷を持つ(塩基性)アミノ酸としては、リジン、アルギニンおよびヒスチジンが挙げられる。負の電荷を持つ(酸性)アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。従って、当然のことながら、アミノ酸は同様の生物物理学的特性を有するアミノ酸で置換することができ、当業者であれば、これらのアミノ酸をコードするヌクレオチド配列を知っているであろう。
【0077】
本明細書(添付の特許請求の範囲、要約書および図面のいずれも含む)に記載されている全ての特徴、および/またはそのように開示されている任意の方法またはプロセスの全ての工程は、そのような特徴および/または工程の少なくとも一部が相互に排他的である組み合わせを除く任意の組み合わせで、上記の態様のいずれかと組み合わせてもよい。
【0078】
本発明のさらなる理解のため、および本発明の実施形態の実施方法を示すために、ここで、例として、添付の図を参照する。
【図面の簡単な説明】
【0079】
【図1】アスパラギン酸が他のアミノ酸に変換される植物生合成経路の一部を示す。関与する酵素は、アスパラギン酸キナーゼ(AK)およびホモセリン不飽和化酵素(HSDH)である。
【図2】アスパラギン酸がトレオニンなどの他のアミノ酸に変換される植物生合成経路を示す。この経路における最初の酵素活性は、アスパラギン酸キナーゼ(AK)である。植物は、二機能性酵素アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)を産生することができる。この生合成経路は、最終産物による正および負のフィードバックを含めて厳密に調節されている。
【図3】シロイヌナズナ中のトレオニンによってアスパラギン酸キナーゼ−ホモセリン脱水素酵素をアロステリック制御するためのモデルを示す(後出のParis et al (2003));
【図4】突然変異型二機能性アスパラギン酸キナーゼ−ホモセリン不飽和化酵素を概略的に示す。
【図5】実施例2において葉片で検出されたトレオニン濃度を示す。
【図6】SAG12プロモーターの配列を示す。
【図7】突然変異型アスパラギン酸キナーゼ−ホモセリン不飽和化酵素の配列を示す。トレオニンによるフィードバック阻害を克服しなければならない突然変異型塩基は、太字でかつ下線が引かれている。
【図8】実施例3に記載されているようなトレオニン非感受性AK−HSDHが老化特異的プロモーターの制御下にある形質転換植物から採取された葉片中のトレオニンの量を示す。
【図9】現場試験により育てられた乾葉中のトレオニン含有量を示す棒グラフである。
【図10】現場試験により育てられた乾葉中のグルタミン含有量を示す棒グラフである。
【図11】現場試験により育てられた乾葉中のグルタミン酸含有量を示す棒グラフである。
【図12】現場試験により育てられた乾葉中のアスパラギン含有量を示す棒グラフである。
【図13】現場試験により育てられた乾葉中のアスパラギン酸含有量を示す棒グラフである。
【図14】現場試験により育てられた乾葉中のヒスチジン含有量を示す棒グラフである。
【図15】本発明に係るベクターの一実施形態のプラスミドマップである。
【図16】本発明に従って使用されるバックボーンベクターのプラスミドマップである。
【発明を実施するための形態】
【実施例】
【0080】
実施例2に記載されているように、本発明者らは、修飾されたフィードバック非感受性アスパラギン酸キナーゼ(AK:HSDH)が葉特異的プロモーターの制御下で発現される形質転換植物を開発した。しかし、本発明者らは、修飾されたAK:HSDHの発現を葉に限定することによって、形質転換植物に対して適応度コストを引き起こすことを見出した。従って、実施例3〜4に記載されるように、本発明者らは次にトレオニン非感受性アスパラギン酸キナーゼ活性(AK:HSDH)を有するポリペプチドをコードするコード配列に連結された老化特異的プロモーター(SAG12)を使用した。得られた形質転換植物は、植物の適応度を損なうことなく、葉の老化時に野生型の濃度よりも高いトレオニンを産生した。
【0081】
実施例1:葉の中のトレオニン濃度に関する試験
緑色の葉または黄変した葉に対してトレオニン濃度に関する試験を行なった。葉片を採取し、分析に使用し、測定は、葉片当たりのトレオニンの量に基づくか(すなわち、thrの量/葉の面積)、上澄み中のタンパク質の量と関連づけられた(すなわち、トレオニンの量/1mgのタンパク質)。葉片を規定の体積の水と一緒にすり潰し、遠心分離して不溶な葉のくずを沈降させた。次いで、このプロセスで得られた上澄みをPhenomenex EZfaast Kitを用いて処理した。これは、標準的なLC/MS装置を用いて定量化することができるように抽出物中のアミノ酸を誘導体化するために使用される専売キットである。
【0082】
較正は、各アミノ酸の定量に関しては外部標準によって行い、誘導体化工程の効率は、内部標準のプロセスに含めることによって試料間で正規化した。クロマトグラムは、ピーク面積で評価し、LC/MSソフトウェアのアルゴリズムにおける積分を用いて濃度と関連づけた。ソフトウェアまたは操作者がピークの特定に確信が持てない場合には、「検出限界未満」と記載される。これは、空のベクター対照試料のいくつかと次世代で分離する無効な(null)植物のいくつかの場合であることが分かった。
【0083】
実施例2:葉特異的プロモーターに作動可能に連結されたフィードバック非感受性AK:HSDHによる形質転換植物
本発明者らは、シロイヌナズナ(At4g19710)由来の二機能性AK:HSDH野生型配列に対して部位特異的突然変異誘発を行なった。従って、最初に下記プライマーを用いるPCRによってシロイヌナズナ由来の葉特異的cDNAライブラリーから野生型配列を単離した:
正方向At4g19710 ATGGCGACTCTGAAGCCGTCATTTAC(配列番号8);
逆方向At4g19710 TTAAGACGGTGCACCGAGATAAGATGC(配列番号9)。
【0084】
この野生型配列を、以下の3つの方法のいずれか1つによって修飾した:(i)AK領域のみを突然変異させるか、(ii)HSDH領域を突然させるか、あるいは(iii)両方の領域を突然変異させて、Thr結合による調節を不可能にした。特異的突然変異は、どちらも酵素調節領域にあるGln443およびGln524に対して行ない、その両方を、アラニンに対する部位特異的突然変異誘発によって突然変異させた(Paris et al (2003), “Mechanism of control of Arabidopsis thaliana aspartate kinase-homoserine dehydrogenase by threonine”, J.Biol.Chem 278:5361-5366およびFrankard et al (1992), “Two feedback-insensitive enzymes of the aspartate pathway in Nicotiana sylvestris” Plant Physiol 99:1285-1293)。
【0085】
この手順のために、Stratagene(登録商標)社製QuikChange(登録商標)部位特異的変異誘発キット(Site-Directed Mutagenesis Kit)(カタログ番号200518)を使用した。部位特異的突然変異誘発反応において、Glu443に対するコドンコードを変えるために、下記プライマーを使用した:
正方向Gln443 GTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTG(配列番号10);および
逆方向Gln443 CAGAATGCTCACTGCTAGCAGCTGATATCATAATTCAC(配列番号11)。
【0086】
Gln524に対して、下記プライマー対を使用した:
正方向Gln524 GTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATG(配列番号12);および
逆方向Gln524 CATTGATCTCAGAACAACCAGCAGATATAGCTCGGAC(配列番号13)。
【0087】
野生型シロイヌナズナAK:HSDHと同様に、これらの3つの突然変異型配列を個別に使用して、タバコ(Nicotiana tabacum)植物を形質転換した。全ての場合に、目的とする遺伝子は、葉特異的エンドウマメ・プラストシアニンプロモーターの制御下で発現された。アグロバクテリウム媒介形質転換を経て全ての個体群の植物を生成し、英国ケンブリッジにおいて温室条件下で育てた。EZfaastアミノ酸キット(Phenomenex(登録商標)社製)を使用して、各試料中の遊離アミノ酸を抽出および誘導体化した。次いで、LC/MSによって定量した。
【0088】
その結果を図5および表1に示す。図5では、AK領域のみが突然変異された一連の植物はAKで始まる名前を有し、HSDH領域のみが突然変異された一連の植物はHSDHで始まる名前を有し、AKおよびHSDH領域の両方が突然変異されている一連の植物はAK/HSDHで始まる名前を有し、野生型植物はWTで始まる名前を有し、空のベクター対照試料を含む形質転換植物はEVで始まる名前を有する。
【0089】
本発明者らは、非突然変異型シロイヌナズナ配列で形質転換された個体群を含む、シロイヌナズナ配列で形質転換された全ての個体群において葉のトレオニン濃度の上昇を達成した。AK領域で突然変異された配列で形質転換された個体群が葉の最も高いThr濃度を示した。これらは、適応度がひどく損なわれたことを示した最も高い割合の個体群と相関していた。本発明者らは、修飾による受精能への影響を抑えるつもりで葉特異的プロモーターを使用したが、それらが受けた影響は、植物全体が酵素に対するフィードバック制御の解除という代謝結果によってもなお影響を受けるには十分であった。
【0090】
しかし、トレオニンの上昇を示した全ての植物において、生育習性の変化との相関性があった。葉は、色が淡く、肥厚し、脆くかつ帯状であった。節間部は短く、成熟度が増すにつれて褐変を示した。芽は発育不良または奇形のいずれかであった。結論として、葉のトレオニンの上昇が得られた点では、部位特異的突然変異誘発は成功であった。しかし、葉に高濃度のトレオニンを有する農学的に生育可能な植物をこの手法から得るつもりであれば、アスパラギン酸キナーゼに対するフィードバック制御の解除による適応度コストをなお克服しなければならない。
【0091】
【表1】
実施例3:老化特異的プロモーターに作動可能に連結されたトレオニン非感受性AK:HSDHを含む形質転換植物
タバコ(Nicotiana tabacum)植物(品種K326)を使用して葉片を得、これを老化特異的プロモーターSAG12(その配列は図6、すなわち配列番号1に示されている)の制御下で、目的の遺伝子(すなわち、突然変異型AK:HSDH、図7(すなわち配列番号2、3および4)に示されているAK:HSDH配列)を保持するバイナリーベクターで(電気穿孔法によって)予め形質転換されていたアグロバクテリウム・ツメファシエンスと共培養した。バイナリーベクターがプロモーターを含むがAK:HSDH遺伝子を含まない対照個体群を同時に育てた。次いで、これらの葉片を組織培養プロトコルによって処理して小植物(Horsch et al. Science 227: 1229-1231, 1985)を得た。各小植物は、細菌からDNAを抽出し、植物のゲノムDNAに組み込む単一の形質転換イベントから得られたものであった。これらの植物を温室に移し、成熟するまで育てた。成熟した葉を植物から切り離し、ポリテン袋に入れて35℃で72時暗所に置いた。この時間の後、この葉を使用して葉片を得、上記実施例1に記載したように、トレオニン含有量を評価した。
【0092】
その結果を図8に示す。図8は、いくつかの植物において高濃度のトレオニンが得られたことを示している。有利なことに、これらの植物は正常に成長した。従って、これらの形質転換植物は、実施例2に記載されている植物で観察されたような適応度コストを示さなかった。
【0093】
実施例4:SAG12プロモーターに連結されたトレオニン非感受性AK:HSDHを含む形質転換植物の現場試験
SAG12:アスパラギン酸キナーゼ(pBNP 138-0253-001)の実験個体群から選択された植物細胞株をノースカロライナで2008年の間現場で育てた。この葉を火力乾燥し、選択した遊離アミノ酸の存在を分析した。この分析は、内部および外部標準較正によって確認されているLC/MSによるものであった。分析物が較正に使用される範囲を超える場合は、表2では「>S」として表中に示し、図では、関連する棒の上に星で示す。
【0094】
この試料一覧を「AK」番号に分類する。この番号は、本発明の遺伝子構築物で修飾されたK326バックグラウンド株、および3種類の代表的な対照試料、すなわち未修飾のK326、未修飾のKV1、未修飾のNC71であり、これら全てを同じ条件下、同じ時間で成長させ、かつ乾燥した。
【0095】
【表2】
図9を参照すると、3種類の対照試料(K326、KV1およびNC7)と比較した5種類の試験植物(AK1〜AK5)によって産生されたトレオニン濃度を示す棒グラフが示されている。図から分かるように、5種類の試験植物全てが、いずれの対照試料よりも著しく多いトレオニンを産生した。さらに、5種類の試験植物はいずれも、それらの成長適応度に対するコストを全く受けなかった。これらのデータは、本発明に係る構築物で形質転換された試験植物が商業用現場条件下で遊離トレオニンが(少なくとも2〜3倍)上昇した葉を生成し、これが乾燥プロセスの間も持続しているという観察を強く支持するものである。さらに、試験植物は、これらの現場条件下において全く不利な点を示さない非分離型であった。
【0096】
また、本発明者らは、現場試験植物から生成された乾葉中のいくつかの他のアミノ酸(GLN:グルタミン;GLU:グルタミン酸;ASN:アスパラギン;ASP:アスパラギン酸;HIS:ヒスチジン)の濃度も評価した。そのデータは図10〜14に示されている。
【0097】
図10から分かるように、細胞株AK3のグルタミン濃度は、3種類の対照細胞株の濃度に匹敵していた。図11に示すように、AK1〜3のグルタミン酸の量は対照試料と同じであったが、AK4〜5の濃度は僅かに高かった。図12は、アスパラギンの各濃度を示す。
【0098】
図13を参照すると、試験細胞株AK1〜AK3のアスパラギン酸濃度は、対照試料とほぼ同じであることが分かるが、AK4〜5の濃度は対照試料よりも高いように思われた。最後に図14を参照すると、ヒスチジン濃度は、試験細胞株AK4が濃度の上昇を示したこと以外は、全体においてほぼ同じであった。
【0099】
要約すると、図10〜14は、これらのアミノ酸の各濃度は対照試料に匹敵するか、対照試料よりもさらに高いことを明らかに示しており、これは、形質転換植物の適応度が損なわれなかったという良い兆候を示している。
【技術分野】
【0001】
本発明は、形質転換植物の産生に使用される遺伝子構築物に関する。この構築物は、特に葉の老化時に、植物に対してそれらの葉の中にトレオニンを蓄積させる能力を持つことができる。本発明は、そのような構築物で形質転換された植物細胞、および形質転換植物そのものに及ぶ。本発明は、形質転換植物の産生方法、および老化した植物中のトレオニン濃度を上昇させる方法にも関する。また、本発明は、遺伝子構築物で形質転換されている収穫された植物の葉(例えば、タバコの葉)、およびそのような収穫された植物の葉を含む喫煙物品にも関する。
【背景技術】
【0002】
火力乾燥されたタバコの風味を高める際の第一の目標は、アミノ酸であるトレオニンの産生である。突然変異型タバコ植物の葉の中に高濃度のトレオニンが蓄積されると、風味および香気上の大きな利点が得られる。しかし、通常は、トレオニンの産生は、アスパラギン酸族の他のアミノ酸、すなわちメチオニン、リジンおよびイソロイシンの産生と共に厳密に調節されている。従って、風味および香気上の利点を得るためには、トレオニンを産生する生合成経路の修飾が必要である。
【0003】
図1および図2に示すように、アスパラギン酸キナーゼ(AK)は、アスパラギン酸をトレオニンなどのアミノ酸に変換する植物生合成経路における最初の酵素である。内在性アスパラギン酸キナーゼは、リジンおよびトレオニンの両方からのフィードバック阻害によって調節されている。従って、フィードバック阻害を受けないアスパラギン酸キナーゼが必要である。しかし、これは、植物を無理にメチオニン飢餓状態したり、アスパラギン酸濃度をそれに依存する他の経路が制限されるまで枯渇させたりしないで達成されなければならない。
【0004】
野生型植物と比較してトレオニンを過剰産生することができる、フィードバック非感受性アスパラギン酸キナーゼを含有する形質転換植物は、成長が不十分であり、かつそのような植物が突然変異に関してホモ接合性であった場合に最悪な適応度コスト(fitness cost)を示すことが以前から分かっている。成長不良および適応度コストは、農業従事者が気付いている植物の成長における有益でない変化である。明らかに、そのような変化はいずれも望ましくない。
【0005】
従って、本発明の発明者らは、上記フィードバック阻害ループを克服するが、理想的にはその適応度に対するいかなるコストも生じさせずに葉の中にトレオニンを蓄積する形質転換植物を提供することを目的とする。これを踏まえて、本発明者らは、トレオニン非感受性アスパラギン酸キナーゼ(AK)酵素をコードする遺伝子がプロモーターの制御下にある複数の遺伝子構築物を開発して、どのような影響があるか、もし影響があれば、この遺伝子の過剰発現が老化した葉の中のトレオニン濃度に影響を与えるか否かを判定した。
【0006】
葉の老化は、葉の細胞が細胞死の前に異なる代謝および構造の変化を受ける植物の発育の一段階である。生理学的および遺伝学的研究では、老化は高度に調節されたプロセスであることが示されている。葉の老化の進行は、葉緑体の分解によって生じるクロロフィルの喪失およびその後の黄変を見かけ上の特徴とする。このような発育段階の特徴である葉のクロロフィル濃度の減少は、例えば、溶媒抽出および分光光度測定によって、あるいは、クロロフィル成分計によって測定することができる。好ましくは一定の条件下で育てられた同じ植物について記録された初期の葉のクロロフィル濃度と比較した葉のクロロフィル濃度の減少は、老化を示す。
【0007】
分子的研究では、老化は遺伝子発現における変化と関連していることが示されている。光合成に関与するタンパク質をコードするmRNAのレベルは老化時に減少するが、老化に関与していると思われているタンパク質をコードする遺伝子のmRNAレベルは増加する。老化は、老化関連遺伝子(SAG)として知られている遺伝子によって調節される高度に組織化されたプロセスである。葉の老化は、タンパク質、核酸および膜の劣化、ならびに、この劣化によってその後に起こる発育中の種子、葉または貯蔵器官などの植物の他の領域への栄養分の輸送と関連する。植物の老化の問題の1つは、老化した葉に存在する多くの有用なミネラルおよび栄養分が葉の中に残留し、そのため、葉が枯れるにつれて実質的に失われることである。老化した葉に存在する多くの他のアミノ酸と同様に、トレオニンは、枯れている葉から取り出さなければ無駄になる。
【0008】
実施例2に記載されているように、本発明者らは、植物中の特定の位置にあるトレオニン非感受性AKを発現する遺伝子構築物を用いる実験、すなわち葉特異的エンドウマメ・プラストシアニンプロモーターを用いる実験を行なった。しかし、発明者らは、そのような形質転換植物の葉は、色が淡く、肥厚し、脆くかつ帯状であることを見出した。節間部は短く、成熟度が増すにつれて褐変を示し、その芽は発育不良または奇形のいずれかであった。従って、そのような形質転換植物は、適応度コストを克服することはできない。
【0009】
そこで、本発明者らは、(SAG12プロモーターの制御下で)植物が老化に差し掛かり、よって正常に発育することが許されている後にのみ、フィードバック非感受性アスパラギン酸キナーゼ(AK)活性が発現される一連の遺伝子構築物を開発した。本発明者らは、フィードバック非感受性アスパラギン酸キナーゼ(AK)が活性化される前に、これらの構築物で形質転換された形質転換植物を成熟(すなわち、老化)まで正常に成長させることができる発明者らが開発した構築物は、意外にも適応度コストを克服することができることに気づいた。
【発明の概要】
【0010】
従って、本発明の第1の態様によれば、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物が提供される。
【0011】
本発明者らは、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に連結された老化特異的プロモーターを使用して第1の態様の構築物を形成し、次いで、これを使用して植物を形質転換させた。本発明者らは研究の結果、意外にも、老化した葉の中のトレオニン濃度が本発明に係る構築物によって上昇することを見出した。さらに、老化特異的プロモーターによって制御される導入遺伝子の発現を一時的に制限することで、トレオニン蓄積植物を産生する初期の試みで以前に認められた適応度コストという負の効果が克服される。実施例に示されているように、得られた形質転換植物は、葉の老化時に野生型の濃度よりも高いトレオニンを産生する。葉の中でトレオニンの蓄積が生じることが実証され、この濃度の上昇が当該構築物を含有するタバコの葉の風味に積極的に寄与すると考えられることから、喫煙物品をそのような葉から製造した。
【0012】
第1の態様の遺伝子構築物におけるプロモーターは、RNAポリメラーゼを誘導して、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に結合させ、かつその転写を開始できるものであればよい。
【0013】
「老化特異的プロモーター(SAG)」は、老化関連遺伝子の発現の制御に関連する任意のプロモーターとすることができる。従って、このプロモーターは、実質的に老化している組織の中でのみ、プロモーターが作動可能に連結されたコード配列(すなわち、遺伝子)の発現を制限することができる。従って、老化特異的プロモーターは、実質的に植物組織が老化している場合にのみ3’タンパク質コード領域の発現が生じるように発生学的に調節されるように、植物組織において遺伝子発現を優先的に促進することができるプロモーターとすることができる。当然のことながら、老化は古い葉などの植物の古い部分で生じ、種子などの植物の若い部分では生じない傾向がある。
【0014】
多数の老化関連遺伝子を発現することで知られている植物の一例は、シロイヌナズナ(Arabidopsis)である。従って、第1の態様に係る構築物に存在するプロモーターは、シロイヌナズナの老化関連遺伝子から単離したものであってもよい。Gepsteinら(The Plant Journal, 2003, 36, 629-642)は、モデルとしてシロイヌナズナを用いて、SAGおよびそのプロモーターについて詳しい研究を行なった。従って、当該遺伝子構築物は、本明細書に開示されている任意のSAG由来のプロモーターを含んでいてもよい。例えば、好適なプロモーターは、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは機能的な断片から選択されてもよい。
【0015】
好ましいプロモーターは、SAG12およびSAG13プロモーターである。一実施形態では、当該プロモーターは、当業者に公知であるSAG12プロモーターまたはその機能的な変異体もしくは断片である(Gan & Amasino, 1997, Plant Physiology, 113: 313-319)。SAG12プロモーターをコードするDNA配列が図6に示されており、本明細書では以下のとおり配列番号1と称する:
配列番号1
TCGAGACCCGATTGTTATTTTTAGACTGAGACAAAAAAGTAGAATCGTTGATTGTTAAAATTTAAAATTAGTTTCATTACGTTTCGATAAAAAAATGATTAGTTTATCATAGCTTAATTATAGCATTGATTTCTAAATTTGTTTTTTGACCACCCTTTTTTCTCTCTTTGGTGTTTTCTTAACATTAGAAGAACCCATAACAATGTACGTTCAAATTAATTAAAAACAATATTTCCAAGTTTTATATACGAAACTTGTTTTTTTTAATGAAAACAGTTGAATAGTTGATTATGAATTAGTTAGATCAATACTCAATATATGATCAATGATGTATATATATGAACTCAGTTGTTATACAAGAAATGAAAATGCTATTTAAATACAGATCATGAAGTGTTAAAAAGTGTCAGAATATGACATGAAGCGTTTTGTCCTACCGGGTATTCGAGTTATAGGTTTGGATCTCTCAAGAATATTTTGGGCCATACTAGTTATATTTGGGCTTAAGCGTTTTGCAAAGAGACGAGGAAGAAAGATTGGGTCAAGTTAACAAAACAGAGACACTCGTATTAGTTGGTACTTTGGTAGCAAGTCGATTTATTTGCCAGTAAAAACTTGGTACACAACTGACAACTCGTATCGTTATTAGTTTGTACTTGGTACCTTTGGTTCAAGAAAAAGTTGATATAGTTAAATCAGTTGTGTTCATGAGGTGATTGTGATTTAATTTGTTGACTAGGGCGATTCCTTCACATCACAATAACAAAGTTTTATAGATTTTTTTTTTATAACATTTTTGCCACGCTTCGTAAAGTTTGGTATTTACACCGCATTTTTCCCTGTACAAGAATTCATATATTATTTATTTATATACTCCAGTTGACAATTATAAGTTTATAACGTTTTTACAATTATTTAAATACCATGTGAAGATCCAAGAATATGTCTTACTTCTTCTTTGTGTAAGAAAACTAACTATATCACTATAATAAAATAATTCTAATCATTATATTTGTAAATATGCAGTTATTTGTCAATTTTGAATTTAGTATTTTAGACGTTATCACTTCAGCCAAATATGATTTGGATTTAAGTCCAAAATGCAATTTCGTACGTATCCCTCTTGTCGTCTAATGATTATTTCAATATTTCTTATATTATCCCTAACTACAGAGCTACATTTATATTGTATTCTAATGACAGGGAAACCTTCATAGAGATTCAGATAGATGAAATTGGTGGGAAACATCATTGAACAGGAAACTTTTAGCAAATCATATCGATTTATCTACAAAAGAATACGTAGCGTAATGAAGTCCACTTGTTGTGAATGACTATGATTTGATCAAATTAGTTAATTTTGTCGAATCATTTTTCTTTTTGATTTGATTAAGCTTTTAACTTGCACGAATGGTTCTCTTGTGAATAAACAGAATCTTTGAATTCAAACTATTTGATTAGTGAAAAGACAAAAGAAGATTCCTTGTTTTTATGTGATTAGTGATTTTGATGCATGAAAGGTACCTACGTACTACAAGAAAAATAAACATGTACGTAACTACGTATCAGCATGTAAAAGTATTTTTTTCCAAATAATTTATACTCATGATAGATTTTTTTTTTTTGAAATGTCAATTAAAAATGCTTTCTTAAATATTAATTTTAATTAATTAAATAAGGAAATATATTTATGCAAAACATCATCAACACATATCCAACTTCGAAAATCTCTATAGTACACAAGTAGAGAAATTAAATTTTACTAGATACAAACTTCCTAATCATCAAATATAAATGTTTACAAAACTAATTAAACCCACCACTAAAATTAACTAAAAATCCGAGCAAAGTGAGTGAACAAGACTTGATTTCAGGTTGATGTAGGACTAAAATGACTACGTATCAAACATCAACGATCATTTAGTTATGTATGAATGAATGTAGTCATTACTTGTAAAACAAAAATGCTTTGATTTGGATCAATCACTTCATGTGAACATTAGCAATTACATCAACCTTATTTTCACTATAAAACCCCATCTCAGTACCCTTCTGAAGTAATCAAATTAAGAGCAAAAGTCATTTAACTTAGG
従って、本発明の構築物におけるプロモーターは、実質的に配列番号1に示されているようなヌクレオチド配列またはその機能的な変異体もしくは機能的な断片を含んでいてもよい。SAG12プロモーター配列は、米国特許第5,689,042号に記載されているようなシロイヌナズナ(Arabidopsis thaliana)から得られたものであってもよい。プロモーターがSAG12である実施形態では、当然のことながら、プロモーターは、配列番号1の1〜2093の各塩基を含んでいてもよい。ただし、プロモーターの機能的な変異体もしくは機能的な断片が本発明の遺伝子構築物に使用されてもよい。
【0016】
「プロモーターの機能的な変異体もしくは機能的な断片」は、そこに作動可能に連結された任意のコード領域の発現を開始させるのに機能的に十分である誘導体またはプロモーターの一部とすることができる。例えば、プロモーターがSAG12に基づいている実施形態では、当業者であれば、配列番号1が修飾されても、SAG12プロモーターの部分のみが必要とされてもよく、従って、当該構築物において、それらがトレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドの遺伝子発現をなお開始させるということを理解しているであろう。同様の修飾を、SAG13、SAG101、SAG21およびSAG18などの他の公知のSAGプロモーターのいずれかのヌクレオチド配列に対して行なってもよい。
【0017】
プロモーターの機能的な変異体および機能的な断片は、転写酵素が推定プロモーター領域に結合し、次いで、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドのコード配列の転写を引き起こすか否かを評価することによって容易に同定することができる。あるいは、そのような機能的な変異体および断片は、コード領域に関連づけられている場合にはプロモーター上で突然変異生成を行ない、遺伝子発現が生じ得るか否かを評価することによって調べてもよい。
【0018】
第1の態様の遺伝子構築物は、老化時にトレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドの発現を引き起こし得るものであればよい。このプロモーターは、トレオニン非感受性アスパラギン酸キナーゼ活性を示すポリペプチドをコードするコード配列の発現を誘導してもよい。従って、当該遺伝子構築物は、トレオニン非感受性アスパラギン酸キナーゼ(AK)をコードする少なくとも1つのコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。従って、第1の実施形態では、遺伝子構築物は、老化特異的プロモーター、およびトレオニン非感受性アスパラギン酸キナーゼ(AK)をコードするコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0019】
実施例3〜4に記載されているように、本発明者らは、本発明の構築物によって植物を形質転換することによって宿主細胞においてトレオニン非感受性アスパラギン酸キナーゼを発現させると、トレオニン濃度の著しい上昇が引き起こされることを見出した。さらに、有利なことに、本発明者らは、本発明の構築物が、形質転換された植物の適応度に対する有害作用を全く有しないことを見出した。
【0020】
図1および図2に示すように、植物では、アミノ酸であるリジン、トレオニン、メチオニンおよびイソロイシンは、アスパラギン酸から合成される。いくつかのフィードバック阻害ループがこの経路で認められている。この経路での最初の酵素であるアスパラギン酸キナーゼ(AK)(EC 2.7.2.4)は、アスパラギン酸のリン酸化を触媒して、付随して生じるATPの加水分解によって3−アスパルチルリン酸を生成する。高等な植物は一般に、少なくとも2種または3種のAKイソ酵素を有すると考えられている。AK活性は、最終産物であるアミノ酸(リジンおよびトレオニン)から負のフィードバックを受ける。少なくとも1種のAKイソ酵素はトレオニンによって、もう1つはリジンおよびS−アデノシルメチオニンによってフィードバック阻害される。オオムギでは、AK活性は、3つのイソ酵素ピーク(isozymal peak)に分類することができ、その内の1つはトレオニンによって阻害され、他の2つはリジンによって阻害される。さらなるフィードバックループは、生合成経路において、ジヒドロピコリン酸合成酵素(DHPS)およびホモセリン不飽和化酵素(HSD)などの他の酵素により作動する。
【0021】
図1および図2に示すように、ホモセリン脱水素酵素(HSDH)としても知られているホモセリン不飽和化酵素(HSD)(EC 1.1.1.3)は、トレオニン、メチオニンおよびイソロイシンの生合成、すなわち3−アスパラギン酸セミアルデヒドのホモセリンへの変換に一意的に関連づけられている第1の反応を触媒する。高等な植物は一般に、少なくとも2つの型のHSDH、すなわちトレオニン感受性型およびトレオニン非感受性型を有する。精製HSDHおよびcDNAクローンの特性評価では、HSDHイソ酵素は単一のタンパク質上のAKに結びづけられていることが確認されており、よって「AK:HSDH」と表わされる。
【0022】
葉のトレオニン(Thr)含有量の上昇は、合成経路への負のフィードバック制御を克服するか否かによって決まる。例えば、大腸菌由来のリジン非感受性AKを発現している形質転換タバコが産生されており、これはトレオニン蓄積表現型を示した。この形質転換体では、細胞質または葉緑体のいずれかを標的にした細菌性AKがタバコにおいて35Sの制御下で発現された。それでも、AKの内因性活性は、リジンおよびトレオニンの両方による阻害をまったく受けなかった。葉緑体を標的とした導入遺伝子は、より高いAK活性を示した。葉緑体型を発現している植物ではより高いトレオニン濃度が認められた。しかし、植物の成長不良が顕著であり、ホモ接合植物は、しわの寄った上葉、開花の遅延および部分的な不稔を含む適応度コストを示した。
【0023】
シロイヌナズナにおいて行われた研究(Paris et al, 2003, The Journal of Biological Chemistry, Vol 278, no. 7, pp5361-5366)では、AK:HSDH酵素の調節領域が、一般的なループ−αへリックス−ループ−βスランド−ループ−βスランドモチーフによって定義される2つの相同サブドメインを含むことが実証されている。部位特異的突然変異誘発が、トレオニン結合部位を解明するために使用された。アスパラギン酸キナーゼ−ホモセリン脱水素酵素のモノマーの各調節領域は、部分的にGln443およびGln524によって構成された2つの非等価トレオニン結合部位を有することが見出された。トレオニンのGln443への結合はAK活性を阻害し、さらに2つ目のトレオニンのGln524への結合を容易にして、それにより、HSDH阻害を引き起こす。
【0024】
図3は、トレオニンによるAK−HSDHの阻害のための提案モデルを示す。ここでは、AKおよびHSDHの活性触媒領域は四角で表わされており、阻害された触媒領域は三角で表わされている。1つ目のサブドメインへのトレオニン(Thr)結合は、(i)他のサブドメインの立体構造変化、および(ii)AK触媒領域の立体構造変化の両方を導き、それによりAK阻害が引き起こされる。2つ目のサブドメインの立体構造変化は、2つ目のトレオニンの結合を誘導し、それによりHSDH触媒領域の立体構造変化およびHSDH阻害が引き起こされる。
【0025】
これらのグルタミン残基のアラニンへの突然変異によって酵素のトレオニン阻害を無効にさせる。この突然変異は、HSDH活性の動態には影響を与えず、トレオニンに対するその感受性のみに影響を与える。すなわち、AK動態は僅かに変化するだけである。しかし、残念なことに、シロイヌナズナ由来のフィードバック非感受性AK:HSDHが導入されかつ発現される形質転換植物は、適応度コストも引き起こす。
【0026】
対照的に、本発明の遺伝子構築物は、老化時にトレオニン非感受性アスパラギン酸活性を有するポリペプチドの発現を引き起こし、形質転換植物の適応度に対する有害作用を全く示さなかった。従って、第1の態様の遺伝子構築物は、トレオニン非感受性アスパラギン酸キナーゼ(AK)、またはトレオニン非感受性の二機能性アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)、あるいはその機能的な変異体もしくは機能的な断片をコードしてもよい。
【0027】
トレオニン非感受性AKまたは二機能性AK−HSDH、あるいはその機能的な変異体もしくは断片は、植物などの任意の好適な供給源に由来するものであってもよい。トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ属、トウモロコシ(Zea)属、フラベリア(Flaveria)属またはクレオメ(Cleome)属に由来するものであってもよい。トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ(Arabidopsis thaliana)、トウモロコシ(Zea mays)、フラベリア・トリネルビア(Flaveria trinervia)、フラベリア・ビデンティス(Flaveria bidentis)、フラベリア・ブラウニー(Flaveria brownie)またはフウチョウソウ(Cleome gynandra)に由来するものであってもよい。好ましくは、当該酵素のコード配列は、シロイヌナズナ(Arabidopsis thaliana)などのシロイヌナズナ(Arabidopsis)属に由来するものであってもよい。
【0028】
特に好ましいトレオニン非感受性酵素は、少なくとも1つのトレオニン結合部位が変化している突然変異型AK−HSDHである。好ましくは、部分的にGln443およびGln524で構成されているトレオニン結合部位の一方または両方が突然変異されている。例えば、シロイヌナズナのAK−HSDHは、Gln443および/またはGln524で突然変異されていてもよい。好ましくは、シロイヌナズナのAK−HSDHは、Gln443およびGln524で突然変異されている。本発明で使用される突然変異型AK:HSDH遺伝子が図7に示されており、ここでは、突然変異型塩基に下線が引かれている。
【0029】
従って、シロイヌナズナのトレオニン非感受性アスパラギン酸キナーゼの一実施形態(すなわち、Gln443Ala単一突然変異体)をコードするDNA配列が、以下のとおり配列番号2として以下に規定される:
配列番号2
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTCARGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号2(すなわち、Q443A)では、強調されているGCTはアラニンをコードする突然変異型Gln443に対応し、強調されているCARは野生型Gln524に対応する。ここで、RはGまたはAのいずれであってもよい。
【0030】
シロイヌナズナトレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln524Ala単一突然変異体)をコードするDNA配列が、以下のとおり配列番号3として以下に規定される:
配列番号3
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCACARGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号3(すなわち、Q524A)では、強調されているGCTはアラニンをコードする突然変異型Gln524に対応し、強調されているCARは野生型Gln443に対応する。ここで、RはGまたはAのいずれであってもよい。
【0031】
シロイヌナズナトレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln443Ala;Gln524Ala二重突然変異体)をコードするDNA配列は、以下のとおり配列番号4として以下に規定される:
配列番号4
ATGGCGACTCTGAAGCCGTCATTTACTGTTTCTCCGCCGAATAGTAATCCGATTAGATTTGGAAGTTTTCCGCCGCAATGCTTTCTCCGTGTTCCGAAACCGCGGCGACTTATATTGCCTAGGTTTCGGAAGACGACTGGTGGTGGCGGCGGCTTGATTCGATGTGAGCTTCCAGATTTTCATCTATCAGCAACAGCAACTACTGTATCAGGTGTATCGACGGTGAATTTAGTGGATCAAGTTCAGATTCCTAAAGGTGAAATGTGGAGTGTTCACAAGTTTGGTGGGACTTGTGTGGGAAACTCTCAGAGGATCAGAAATGTAGCAGAGGTTATAATCAATGATAATTCCGAAAGAAAACTTGTGGTTGTCTCGGCGATGTCGAAGGTTACGGACATGATGTATGACTTAATCCGCAAGGCACAATCACGAGATGATTCTTATTTATCCGCGTTGGAAGCTGTCTTGGAAAAGCATCGTTTAACAGCTCGTGACCTTCTCGATGGAGATGATCTCGCTAGTTTCTTGTCACATTTGCATAATGATATTAGTAATCTTAAAGCAATGCTTCGTGCTATATACATAGCTGGCCATGCATCAGAGTCGTTTTCAGATTTTGTTGCAGGACATGGGGAGCTTTGGTCTGCTCAGATGCTATCATATGTTGTCAGAAAGACTGGGCTTGAGTGCAAGTGGATGGATACTAGAGACGTGCTCATTGTTAATCCCACCAGCTCTAATCAGGTTGATCCTGATTTTGGTGAATCTGAGAAGAGACTCGATAAATGGTTCTCCTTAAATCCGTCGAAAATTATTATTGCGACTGGGTTTATTGCTAGCACTCCGCAAAATATTCCAACAACTTTGAAAAGAGATGGGAGTGATTTCTCAGCAGCTATTATGGGTGCTTTATTGAGAGCTCGTCAAGTAACCATTTGGACAGATGTTGATGGTGTATACAGTGCGGATCCTCGTAAAGTTAATGAGGCAGTGATACTCCAGACACTTTCTTATCAAGAGGCCTGGGAAATGTCTTATTTTGGAGCAAATGTGTTACATCCTCGCACCATCATTCCTGTGATGCGATATAATATTCCGATTGTGATTAGAAATATTTTCAATCTCTCTGCACCGGGAACAATAATCTGTCAACCTCCTGAAGATGATTATGACCTTAAACTGACAACTCCTGTCAAAGGGTTTGCAACTATTGACAATTTGGCCCTCATAAATGTTGAAGGTACTGGAATGGCTGGTGTACCCGGTACTGCAAGTGACATTTTTGGCTGTGTAAAAGATGTTGGAGCTAATGTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTGTGTGCTTTGCTGTGCCTGAGAAGGAAGTAAACGCAGTCTCTGAGGCATTGCGGTCGAGATTTAGTGAAGCTTTACAAGCGGGACGTCTTTCTCAGATTGAGGTGATACCAAACTGTAGCATCTTAGCTGCAGTCGGCCAGAAAATGGCTAGTACACCTGGAGTTAGTTGTACACTTTTCAGTGCTTTGGCGAAGGCTAATATTAATGTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATGTTACTGTCGTTATTAAACGTGAAGATAGCGTTAAGGCGTTAAGAGCTGTACACTCGAGGTTTTTCTTGTCAAGAACAACATTAGCAATGGGAATCGTAGGACCGGGCTTGATTGGTGCAACATTACTTGACCAGCTGCGGGATCAGGCTGCTGTTCTCAAACAAGAATTTAACATTGATCTGCGTGTTTTGGGAATCACTGGTTCAAAGAAGATGTTATTGAGTGACATTGGTATTGATTTGTCGAGATGGAGAGAACTTCTAAACGAGAAGGGAACAGAGGCGGATTTGGATAAATTCACTCAACAAGTGCATGGAAATCATTTTATCCCCAACTCTGTAGTGGTTGATTGTACAGCAGACTCTGCTATTGCAAGCCGTTACTATGATTGGTTACGAAAGGGAATTCATGTCATTACCCCAAATAAAAAGGCTAACTCAGGTCCCCTCGATCAGTACTTGAAACTGAGAGATCTTCAAAGGAAATCCTACACTCATTACTTCTACGAAGCCACTGTTGGAGCTGGTCTTCCAATTATCAGCACTTTACGTGGTCTCCTTGAGACAGGAGATAAGATACTACGCATAGAGGGCATTTGCAGTGGAACTTTGAGTTATCTATTCAACAATTTTGTTGGAGATCGAAGTTTCAGCGAGGTTGTCACTGAAGCAAAGAACGCAGGTTTCACTGAGCCTGATCCAAGAGATGATTTATCTGGAACTGATGTTGCAAGGAAGGTGATTATCCTCGCTCGAGAATCTGGACTGAAATTGGACCTCGCTGATCTCCCCATTAGAAGTCTCGTACCAGAACCTCTAAAAGGATGCACTTCTGTTGAAGAATTCATGGAGAAACTCCCACAGTACGATGGAGACCTAGCAAAAGAAAGGCTAGATGCTGAAAACTCTGGGGAAGTTCTGAGATATGTTGGAGTGGTGGACGCTGTTAACCAAAAGGGAACAGTTGAACTTCGAAGATACAAGAAAGAACATCCATTTGCGCAGCTCGCAGGTTCAGACAACATAATAGCCTTCACAACGACAAGGTACAAGGATCATCCACTTATAGTCCGAGGACCTGGAGCTGGTGCTCAAGTCACGGCCGGTGGTATATTCAGCGACATACTAAGGCTTGCATCTTATCTCGGTGCACCGTCTTAA
配列番号4(すなわち、Q443A;Q524A)では、強調されているGCTは、どちらもアラニンをコードする突然変異型Gln443およびGln524に対応する。
【0032】
従って、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列は、実質的に配列番号2、3または4のうちのいずれか1つに示されているような核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0033】
トレオニン非感受性アスパラギン酸キナーゼの一実施形態(すなわち、Gln443Ala単一突然変異体)のポリペプチド配列が、以下のとおり配列番号5として以下に規定される:
配列番号5
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISAASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISQGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGG
IFSDILRLASYLGAPS 配列番号5では、位置443にある突然変異型アラニン(A)、および位置524にある野生型グルタミン(Q)が強調されている。
【0034】
トレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln524Ala単一突然変異体)のポリペプチド配列は、以下のとおり配列番号6として以下に規定される:
配列番号6
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISQASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISAGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGGIFSDILRLASYLGAPS
配列番号6では、位置524にある突然変異型アラニン(A)および位置443にある野生型グルタミン(Q)が強調されている。
【0035】
トレオニン非感受性アスパラギン酸キナーゼの別の実施形態(すなわち、Gln443Ala;Gln524Ala二重突然変異体)のポリペプチド配列は、以下のとおり配列番号7として以下に規定される:
配列番号7
MATLKPSFTVSPPNSNPIRFGSFPPQCFLRVPKPRRLILPRFRKTTGGGGGLIRCELPDF
HLSATATTVSGVSTVNLVDQVQIPKGEMWSVHKFGGTCVGNSQRIRNVAEVIINDNSERK
LVVVSAMSKVTDMMYDLIRKAQSRDDSYLSALEAVLEKHRLTARDLLDGDDLASFLSHLH
NDISNLKAMLRAIYIAGHASESFSDFVAGHGELWSAQMLSYVVRKTGLECKWMDTRDVLI
VNPTSSNQVDPDFGESEKRLDKWFSLNPSKIIIATGFIASTPQNIPTTLKRDGSDFSAAI
MGALLRARQVTIWTDVDGVYSADPRKVNEAVILQTLSYQEAWEMSYFGANVLHPRTIIPV
MRYNIPIVIRNIFNLSAPGTIICQPPEDDYDLKLTTPVKGFATIDNLALINVEGTGMAGV
PGTASDIFGCVKDVGANVIMISAASSEHSVCFAVPEKEVNAVSEALRSRFSEALQAGRLS
QIEVIPNCSILAAVGQKMASTPGVSCTLFSALAKANINVRAISAGCSEYNVTVVIKREDS
VKALRAVHSRFFLSRTTLAMGIVGPGLIGATLLDQLRDQAAVLKQEFNIDLRVLGITGSK
KMLLSDIGIDLSRWRELLNEKGTEADLDKFTQQVHGNHFIPNSVVVDCTADSAIASRYYD
WLRKGIHVITPNKKANSGPLDQYLKLRDLQRKSYTHYFYEATVGAGLPIISTLRGLLETG
DKILRIEGICSGTLSYLFNNFVGDRSFSEVVTEAKNAGFTEPDPRDDLSGTDVARKVIIL
ARESGLKLDLADLPIRSLVPEPLKGCTSVEEFMEKLPQYDGDLAKERLDAENSGEVLRYV
GVVDAVNQKGTVELRRYKKEHPFAQLAGSDNIIAFTTTRYKDHPLIVRGPGAGAQVTAGGIFSDILRLASYLGAPS
配列番号7では、位置443および524にある突然変異型アラニン(A)が強調されている。
【0036】
従って、トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチド配列は、実質的に配列番号5、6または7のうちのいずれか1つに示されているようなアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0037】
本発明の遺伝子構築物は、発現カセットの形態であってもよく、宿主細胞におけるコード配列の発現に適したものであればよい。本発明の遺伝子構築物は、それがベクターに組み込まれることなく、宿主細胞に導入されてもよい。例えば、核酸分子であり得る遺伝子構築物は、リポソームまたはウイルス粒子内に組み込まれてもよい。あるいは、精製された核酸分子(例えば、ヒストンを含まないDNA、すなわち裸のDNA)が、例えば、直接的なエンドサイトーシス取り込みなどの好適な手段によって宿主細胞に直接挿入されてもよい。当該遺伝子構築物は、形質移入、感染、微量注入、細胞融合、原形質融合または衝撃照射(ballistic bombardment)によって宿主対象(例えば、植物)の細胞に直接導入されてもよい。あるいは、本発明の遺伝子構築物は、パーティクルガン法を用いて宿主細胞に直接導入されてもよい。あるいは、当該遺伝子構築物は、好適な宿主細胞における発現のために組換え型ベクター内部に含まれていてもよい。
【0038】
従って、第2の態様では、第1の態様に係る遺伝子構築物を含む組換え型ベクターが提供される。
【0039】
当該組換え型ベクターは、プラスミド、コスミドまたはファージであってもよい。そのような組換え型ベクターは、宿主細胞を第1の態様の遺伝子構築物で形質転換し、かつその中の発現カセットを複製するのに非常に有用である。当業者であれば、本発明の遺伝子構築物は、発現のために、多くの種類のバックボーンベクターと組み合わせ得ることを理解しているであろう。バックボーンベクターは、例えば、1つのベクターが大腸菌およびアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)の両方で複製することができるバイナリーベクターであってもよい。例えば、好適なベクターは、pBIN19などのpBINプラスミドであってもよい。しかし、好ましいバックボーンベクターはBNP1380000001であり、これは、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290) に基づいており、かつSAG12プロモーターを内部に含む。このベクターの一実施形態が図16に示されている。
【0040】
組換え型ベクターは、プロモーター(例えば、老化関連プロモーター)に加えて、様々な他の機能的な成分、および少なくとも1つの(突然変異型AK−HSDHをコードする)コード配列を含んでいてもよい。例えば、組換え型ベクターは、宿主細胞中の細胞質ゾル内で自己複製するように設計されていてもよい。この場合、組換え型ベクター内に、DNA複製を誘導または調節する成分が必要な場合がある。あるいは、組換え型ベクターは、宿主細胞のゲノムに組み込まれるように設計されていてもよい。この場合、(例えば相同組換えによる)標的組換えを支持するDNA配列が想定される。
【0041】
また、当該組換え型ベクターは、クローニングプロセスにおける選択可能なマーカーとして、すなわち、形質移入または形質転換されている細胞の選択を可能にするため、および異種のDNAが組み込まれているベクターを内部に含む細胞の選択を可能にするために使用できる遺伝子のDNAコードを含んでいてもよい。あるいは、選択可能なマーカー遺伝子は、目的の遺伝子を含有するベクターと同時に使用される異なるベクター中に存在していてもよい。当該ベクターは、コード配列の発現の調節に関与するDNA、または発現されるポリペプチドをある種の宿主細胞、例えば葉緑体に導くためのDNAを含んでいてもよい。従って、第2の態様のベクターは、選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子)、ポリペプチド終結シグナルおよびタンパク質標的配列(例えば、葉緑体輸送ペプチド)からなる群から選択される少なくとも1つのさらなる成分を含んでいてもよい。
【0042】
好適なマーカー遺伝子の例としては、抗生物質耐性遺伝子、例えば、カナマイシン、ジェネテシン(G418)およびハイグロマイシン(npt−II、hyg−B)に対して耐性を示す遺伝子;除草剤耐性遺伝子、例えば、ホスフィノトリシンおよびスルホンアミド系除草剤(それぞれbarおよびsuI;欧州特許出願公開第242246号、欧州特許出願公開第0249637号)に対して耐性を示す遺伝子;およびスクリーニング可能なマーカー、例えば、β−グルクロニダーゼ(英国特許出願公開第2197653号)、ルシフェラーゼおよび緑色蛍光タンパク質(GFP)が挙げられる。
【0043】
当該マーカー遺伝子は、第2のプロモーター(老化関連プロモーターであってもそうでなくてもよい)によって制御されてもよく、細胞における発現を可能にし、種子の中に存在するものであってもそうでなくてもよく、それにより、植物の発育の任意の段階でマーカーを含有する細胞または組織の選択を可能にする。好適な第2のプロモーターは、アグロバクテリウム(Agrobacterium)のノパリン合成酵素遺伝子のプロモーターおよび35Sカリフラワーモザイクウイルス(CaMV)転写物をコードする遺伝子に由来するプロモーターである。ただし、任意の他の好適な第2のプロモーターが使用されてもよい。
【0044】
本発明の遺伝子構築物の様々な実施形態は、実施例2に記載されていて、かつ以下のように要約することができる好適なクローニング法を用いて産生されてもよい。野生型AK−HSDHをコードする遺伝子は、好適なプライマーを用いるPCRによってゲノムまたはcDNAの鋳型のいずれかから増幅されてもよい。野生型AK−HSDH遺伝子の増幅に適したプライマーは、配列番号8および/または配列番号9であってもよい。PCR産物は、アガロースゲル電気泳動法を用いて調べてもよい。次いで、位置443および/または524にある野生型コドンを突然変異させてGln443AlaおよびGln524Ala単一突然変異体または二重突然変異体を産生するために、好適な対のプライマーを用いる部位特異的突然変異誘発を行なってもよい。例えば、Gln443のコドンを変化させるのに適したプライマーは、配列番号10および/または配列番号11であってもよい。Gln524のコドンを変化させるのに適したプライマーは、配列番号12および/または配列番号13であってもよい。
【0045】
次いで、この2つの単一突然変異体または二重突然変異体のうちのいずれか一方をコードするPCR産物を、クローニングに適したベクター、例えば、Invitrogen社から入手でき、pCR4 Blunt-TOPOベクターという商品名で市販されているベクターの中に連結してもよい。次いで、PCR産物を内部に含むベクターを大腸菌などの好適な宿主中で成長させてもよい。次いで、大腸菌コロニーを、好適なプライマーを用いるPCRによってスクリーニングしてもよく、正しい制限酵素消化パターンを示すプラスミド中の挿入断片を好適なプライマーを用いて配列決定してもよい。
【0046】
TOPO−cDNA(AK−HSDH)を保持する大腸菌コロニーを培養して好適な量のプラスミドを作製し、次いで、これを精製してもよい。ついで、プラスミドを温浸して、突然変異型AK−HSDHをコードするDNA断片を遊離させた後、これを、pBNPプラスミドなどの、好適なプロモーター(例えば、SAGプロモーター(好ましくは、SAG12))を内部に含むベクターにクローン化してもよい。得られたプラスミドは、pBNP138−0453−001と命名された。第2の態様に係るベクターの一実施形態は、実質的に図15に示されているようなものであってもよい。
【0047】
第3の態様では、同じ条件下で栽培された対応する野生型植物よりも高濃度のトレオニンを葉に蓄積させる形質転換植物の産生方法が提供される。当該方法は、
(i)植物細胞を、第1の態様の遺伝子構築物または第2の態様のベクターで形質転換することと、
(ii)形質転換された細胞から植物を再生することと、
を含む。
【0048】
植物の葉の中のトレオニン濃度および植物の成長率の測定方法が、実施例1に記載されている。第3の態様の方法は、試験植物細胞を第1の態様に係る遺伝子構築物または第2の態様に係るベクターで形質転換することを含む。当該遺伝子構築物またはベクターは、任意の好適な手段によって宿主細胞に導入してもよい。
【0049】
第4の態様では、第1の態様に係る遺伝子構築物または第2の態様に係る組換え型ベクターを含む細胞が提供される。
【0050】
当該細胞は植物細胞であってもよい。本発明者らは、宿主細胞における老化特異的プロモーターの制御下でのトレオニン非感受性アスパラギン酸キナーゼの発現は、意外にも、適応度を損なうことなく老化した葉の中のトレオニン濃度を上昇させるのに有効であることに気づいたため、第4の態様の細胞は、1つまたは複数の第1の態様の構築物あるいは1つまたは複数の第2の態様のベクターを含んでいてもよい。
【0051】
当該細胞は、公知の技術を用いて本発明に係る遺伝子構築物またはベクターで形質転換されていてもよい。遺伝子構築物を宿主細胞に導入するのに適した手段は、例えば、欧州特許出願公開第0116718号および第0270822号に記載されているように当該技術分野で知られている手段による、アグロバクテリウムが保持するディスアーム型Tiプラスミドベクターの使用を含んでもよい。さらなる方法は、植物原形質を形質転換することであってもよく、この方法では、最初に細胞壁を除去し、核酸を導入し、次いで、細胞壁を再編成する。次いで、形質転換された細胞を植物に成長させてもよい。
【0052】
第5の態様では、第1の態様に係る遺伝子構築物または第2の態様に係るベクターを含む形質転換植物が提供される。
【0053】
第5の態様に係る形質転換植物としては、アブラナ属などのアブラナ科が挙げられる。当該植物は、セイヨウアブラナ(Brassica napus)(菜種)であってもよい。
【0054】
第5の態様に係る形質転換植物のさらなる例としては、コムギ連(Triticeae)などのイネ科が挙げられる。当該植物は、コムギ属(Triticum)(コムギ)であってもよい。コムギ中の種子タンパク質含有量を増加させることによって、パンなどのそのようなコムギを含む食品の体積を増加させてもよい。
【0055】
第5の態様に係る好適な形質転換植物のさらなる例としては、植物のナス科が挙げられ、ナス科の例としては、例えば、チョウセンアサガオ、ナス、マンドレイク、ベラドンナ(deadly nightshade (belladonna))、トウガラシ(パプリカ、カラトウガラシ)、ジャガイモおよびタバコが挙げられる。ナス科の好適な属の一例はニコチアナである。ニコチアナの好適な種は、タバコ植物、または単にタバコと称される場合がある。植物を第1の態様の遺伝子構築物または第2の態様のベクターで形質転換する様々な方法が知られており、本発明で使用することができる。
【0056】
例えば、タバコは、以下のように形質転換してもよい。タバコ(Nicotiana tabacum)を、基本的にHorsch et al. (Science 227: 1229-1231, 1985)に記載されているように葉片共培養法を用いて形質転換した。最も若い2枚の広がった葉を7週齢のタバコ植物から採取してもよく、8%のドメストス(商標)中で10分間表面滅菌し、滅菌蒸留水で6回洗浄してもよい。葉片をNo.6コルク穿孔器を用いて切断し、約2分間かけて(本発明に係る)適当なバイナリーベクターを含有するアグロバクテリウム懸濁液に入れてもよい。この葉片を、2枚の無菌濾紙の間に穏やかにブロットしてもよい。10枚の葉片をLS3%スクロース+2μMのBAP+0.2μMのNAAを含むプレート上に置き、次いで、これを成長室(growth room)において2日間インキュベートしてもよい。
【0057】
葉片は、クラフォラン500g/lおよびのカナマイシン100g/lを添加したLS+3%スクロース+2μMのBAP+0.2μMのNAAを含むプレートに移してもよい。この葉片は、2週間後に上記培地を含む新しいプレートに移してもよい。さらに2週間後、この葉片は、クラフォラン500mg/lおよびカナマイシン100mg/lを添加したLS+3%スクロース+0.5μMのBAPを含むプレートに移してもよい。この葉片は、2週間ごとに新しい培地に移してもよい。苗条が現れたら、それらを切除し、クラフォラン500mg/lを添加したLS+3%スクロースを入れた瓶に移してもよい。瓶の中の苗条は、約4週間後、LS+3%スクロース+250mg/lのクラフォランに移してもよい。さらに3〜4週間後、この植物を、LS+3%スクロース(抗生物質なし)に移して、根づかせてもい。植物が根づいたら、温室の土壌に移してもよい。
【0058】
第6の態様では、第5の態様に係る形質転換植物から得られた植物の繁殖用製品(propagation product)が提供される。
【0059】
「植物繁殖用製品」は、そこからさらなる植物が生産することができる任意の植物とすることができる。好適には、この植物繁殖用製品は種子であってもよい。
【0060】
また、本発明は、同じ条件下で栽培された対応する野生型植物から収穫された葉よりも高濃度のトレオニンを含有する、本発明の形質転換植物から収穫された葉も包含する。
【0061】
従って、第7の態様では、同じ条件下で栽培された野生型植物から採取された、収穫された葉の中の対応するトレオニン濃度よりも高濃度のトレオニンを含有する収穫された葉が提供され、ここで、この葉は、第5の態様に係る形質転換植物から収穫されたものか、第3の態様に係る方法によって生産されたものである。
【0062】
本発明の第8の態様は、老化した葉の中でトレオニンを過剰産出することができる突然変異型タバコ植物から得られたトレオニンが豊富なタバコを含む喫煙物品を提供する。
【0063】
有利かつ好ましくは、突然変異型タバコ植物は、本発明の遺伝子構築物またはベクターで形質転換されたものであってもよい。当該喫煙物品は、紙巻タバコ、葉巻タバコ、シガリロ(小型葉巻)または手巻きタバコなどであってもよい。
【0064】
トレオニンが減少したタバコには、同じ条件下で栽培された野生型植物中の対応する濃度よりもトレオニン濃度の高いタバコが含まれる。そのような喫煙物品は、突然変異型タバコ植物から得られたタバコを含んでいてもよく、このタバコは、本発明の第1の態様に係る遺伝子構築物または第2の態様に係るベクターで形質転換されたものであってもよい。トレオニンが豊富なタバコは風味および香気がよくなっている。
【0065】
当然のことながら、本発明は、植物の適応度を損なうことなく、対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、植物の代謝を変化させて、葉の老化開始後にトレオニンの産生の増加を達成することを含む方法を提供する。
【0066】
従って、本発明の第9の態様では、植物の適応度を損なうことなく、対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、葉の老化開始後にトレオニンの産生の増加を達成することを含む方法が提供される。
【0067】
好ましくかつ有利なことには、本発明に係る方法は、生産される植物の健康または適応度を損なうことはない。好ましくは、当該方法は、第1の態様の遺伝子構築物または第2の態様のベクターを用いて、試験植物および好ましくはその葉を形質転換することを含む。
【0068】
実施例4に記載されているように、本発明の形質転換された植物中のトレオニン濃度を測定し、かつ老化した葉の中のトレオニン濃度の上昇を示すことに加えて、本発明者らは、形質転換植物中のグルタミン、グルタミン酸、アスパラギン酸およびヒスチジンなどの他のアミノ酸の濃度も測定した。図10〜図14に示すように、これらの各アミノ酸の濃度は、対照試料に匹敵するかそれのよりも高く、これは、形質転換植物の適応度が損なわれていなかったことを強く示している。従って、要約すると、本発明の構築物は、形質転換植物の適応度に悪影響を与えることなく、老化した葉の中のトレオニン濃度を上昇させるために好適に使用することができる。
【0069】
当然のことながら、本発明は、実質的に本明細書で参照されている配列のいずれかのアミノ酸または核酸配列(その機能的な変異体もしくは機能的な断片を含む)を含む、任意の核酸、ペプチド、その変異体、誘導体または類似体にまで及ぶ。「実質的にアミノ酸/ポリヌクレオチド/ポリペプチド配列」、「機能的な変異体」および「機能的な断片」という用語は、本明細書で参照されている配列のうちのいずれか1つのアミノ酸/ポリヌクレオチド/ポリペプチド配列との少なくとも40%の配列同一性、例えば、配列番号1(すなわちSAG12プロモーター)として同定されているプロモーターまたは(AK−HSDH酵素の様々な実施形態をコードする)配列番号2、3または4として同定されている遺伝子との40%の同一性、あるいは配列番号5、6または7(すなわち、突然変異型AK−HSDH酵素の様々な実施形態)として同定されているポリペプチドとの40%の同一性などを有する配列とすることができる。
【0070】
また、参照されている配列のいずれかに対して65%を超える、より好ましくは70%を超える、さらにより好ましくは75%を超える、なおより好ましくは80%を超える配列の同一性を有するアミノ酸/ポリヌクレオチド/ポリペプチド配列も想定される。好ましくは、アミノ酸/ポリヌクレオチド/ポリペプチド配列は、参照されている配列のいずれかとの少なくとも85%の同一性、より好ましくは少なくとも90%の同一性、さらにより好ましくは少なくとも92%の同一性、なおより好ましくは少なくとも95%の同一性、なおより好ましくは少なくとも97%の同一性、なおより好ましくは少なくとも98%の同一性、最も好ましくは本明細書で参照されている配列のいずれかとの少なくとも99%の同一性を有する。
【0071】
当業者であれば、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算方法が分かっているであろう。2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合を計算するために、最初に2つの配列のアラインメントを作成した後、配列の同一性値を計算しなければならない。2つの配列の同一性の割合は、(i)配列を整列させるために使用される方法、例えば、ClustalW、BLAST、FASTA、Smith−Waterman法(異なるプログラムにより実行される)、あるいは3D比較による構造アラインメント、および(ii)アラインメント法(例えば、ローカルまたはグローバルアラインメント)によって使用されるパラメータ、使用される対のスコア行列(例えば、BLOSUM62、PAM250、Gonnetなど)、およびギャップペナルティ(例えば、関数および定数)によって異なる値を取る可能性がある。
【0072】
アラインメント作成後の、2つの配列間の同一性の割合に関する計算方法は数多くある。例えば、(i)最も短い配列の長さ、(ii)アラインメントの長さ、(iii)配列の平均長、(iv)ギャップのない位置の数、または(iv)オーバーハングを除く等価位置の数によって同一部分の数を割ってもよい。さらに、当然のことながら、同一性の割合は長さにも強く依存する。従って、一対の配列が短い程、偶然に生じることを予測し得る配列の同一性が高くなる。
【0073】
従って、当然のことながら、タンパク質またはDNA配列の正確なアラインメントは、複雑なプロセスである。有名なマルチプルアライメントプログラムであるClustalW(Thompson et al., 1994, Nucleic Acids Research, 22, 4673-4680; Thompson et al., 1997, Nucleic Acids Research, 24, 4876-4882)は、本発明に従ってタンパク質またはDNAのマルチプルアラインメントを生成する好ましい方法である。ClustalWの好適なパラメータは、以下のとおりであってもよい:DNAアラインメントについては:ギャップ開始ペナルティー(Gap Open Penalty)=15.0、ギャップ伸長ペナルティー(Gap Extension Penalty)=6.66および行列=同一性。タンパク質アラインメントについては:ギャップ開始ペナルティー=10.0、ギャップ伸長ペナルティー=0.2および行列=Gonnet。DNAおよびタンパク質アラインメントについては:ENDGAP=−1およびGAPDIST=4。当業者であれば、最適な配列アラインメントのためにこれらのパラメータおよび他のパラメータを変える必要があるかも知れないことが分かっているであろう。
【0074】
好ましくは、次に、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算は、(N/T)*100(式中、Nは、同一の残基を共有する配列の位置の数であり、Tは、ギャップを含めるがオーバーハングを除いて比較される位置の合計数である)としてそのようなアラインメントから計算してもよい。従って、2つの配列間の同一性の割合の最も好ましい計算方法は、(i)好適な組み合わせのパラメータ(例えば、上記のようなパラメータ)を用いるClustalWプログラムを用いて配列アラインメントを作成すること、および(ii)NおよびTの値を式:配列の同一性=(N/T)*100に書き込むことを含む。
【0075】
同様の配列の他の同定方法が当該技術分野で知られている。例えば、実質的に同様のヌクレオチド配列は、厳密な条件下で、配列番号1、2、3または4に示されている配列あるいはそれらの相補体とハイブリッドを形成する配列によってコードされるであろう。厳密な条件とは、ヌクレオチドが約45℃で3倍の塩化ナトリウム/クエン酸ナトリウム(SSC)中でフィルター結合DNAまたはRNAとハイブリッド形成した後、約20〜65℃で0.2倍のSSC/0.1%SDS中で少なくとも1回洗浄されることを意味する。あるいは、実質的に同様のポリペプチドは、配列番号5、6または7に示されている配列とは、少なくとも1個、ただし5、10、20、50または100個未満のアミノ酸が異なっていてもよい。
【0076】
遺伝子コードの縮重により、本明細書に記載されている任意の核酸配列を、それによってコードされるタンパク質の配列に実質的に影響を与えることなく変化または変更させてその機能的な変異体を得ることができることは明らかである。好適なヌクレオチド変異体は、配列内の同じアミノ酸をコードする異なるコドンの置換によって静的な変化(silent change)を生じさせることにより変化した配列を有する変異体である。他の好適な変異体は、相同ヌクレオチド配列を有するが、置換されたアミノ酸と同様の生物物理学的特性の側鎖を有するアミノ酸をコードする異なるコドンの置換によって保存的変化(conservative change)を生じさせることにより変化した配列の全てもしくは部分を含む変異体である。例えば、小型で非極性の疎水性アミノ酸としては、グリシン、アラニン、ロイシン、イソロイシン、バリン、プロリンおよびメチオニンが挙げられる。大型で非極性の疎水性アミノ酸としては、フェニルアラニン、トリプトファンおよびチロシンが挙げられる。中立極性のアミノ酸としては、セリン、トレオニン、システイン、アスパラギンおよびグルタミンが挙げられる。正電荷を持つ(塩基性)アミノ酸としては、リジン、アルギニンおよびヒスチジンが挙げられる。負の電荷を持つ(酸性)アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。従って、当然のことながら、アミノ酸は同様の生物物理学的特性を有するアミノ酸で置換することができ、当業者であれば、これらのアミノ酸をコードするヌクレオチド配列を知っているであろう。
【0077】
本明細書(添付の特許請求の範囲、要約書および図面のいずれも含む)に記載されている全ての特徴、および/またはそのように開示されている任意の方法またはプロセスの全ての工程は、そのような特徴および/または工程の少なくとも一部が相互に排他的である組み合わせを除く任意の組み合わせで、上記の態様のいずれかと組み合わせてもよい。
【0078】
本発明のさらなる理解のため、および本発明の実施形態の実施方法を示すために、ここで、例として、添付の図を参照する。
【図面の簡単な説明】
【0079】
【図1】アスパラギン酸が他のアミノ酸に変換される植物生合成経路の一部を示す。関与する酵素は、アスパラギン酸キナーゼ(AK)およびホモセリン不飽和化酵素(HSDH)である。
【図2】アスパラギン酸がトレオニンなどの他のアミノ酸に変換される植物生合成経路を示す。この経路における最初の酵素活性は、アスパラギン酸キナーゼ(AK)である。植物は、二機能性酵素アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)を産生することができる。この生合成経路は、最終産物による正および負のフィードバックを含めて厳密に調節されている。
【図3】シロイヌナズナ中のトレオニンによってアスパラギン酸キナーゼ−ホモセリン脱水素酵素をアロステリック制御するためのモデルを示す(後出のParis et al (2003));
【図4】突然変異型二機能性アスパラギン酸キナーゼ−ホモセリン不飽和化酵素を概略的に示す。
【図5】実施例2において葉片で検出されたトレオニン濃度を示す。
【図6】SAG12プロモーターの配列を示す。
【図7】突然変異型アスパラギン酸キナーゼ−ホモセリン不飽和化酵素の配列を示す。トレオニンによるフィードバック阻害を克服しなければならない突然変異型塩基は、太字でかつ下線が引かれている。
【図8】実施例3に記載されているようなトレオニン非感受性AK−HSDHが老化特異的プロモーターの制御下にある形質転換植物から採取された葉片中のトレオニンの量を示す。
【図9】現場試験により育てられた乾葉中のトレオニン含有量を示す棒グラフである。
【図10】現場試験により育てられた乾葉中のグルタミン含有量を示す棒グラフである。
【図11】現場試験により育てられた乾葉中のグルタミン酸含有量を示す棒グラフである。
【図12】現場試験により育てられた乾葉中のアスパラギン含有量を示す棒グラフである。
【図13】現場試験により育てられた乾葉中のアスパラギン酸含有量を示す棒グラフである。
【図14】現場試験により育てられた乾葉中のヒスチジン含有量を示す棒グラフである。
【図15】本発明に係るベクターの一実施形態のプラスミドマップである。
【図16】本発明に従って使用されるバックボーンベクターのプラスミドマップである。
【発明を実施するための形態】
【実施例】
【0080】
実施例2に記載されているように、本発明者らは、修飾されたフィードバック非感受性アスパラギン酸キナーゼ(AK:HSDH)が葉特異的プロモーターの制御下で発現される形質転換植物を開発した。しかし、本発明者らは、修飾されたAK:HSDHの発現を葉に限定することによって、形質転換植物に対して適応度コストを引き起こすことを見出した。従って、実施例3〜4に記載されるように、本発明者らは次にトレオニン非感受性アスパラギン酸キナーゼ活性(AK:HSDH)を有するポリペプチドをコードするコード配列に連結された老化特異的プロモーター(SAG12)を使用した。得られた形質転換植物は、植物の適応度を損なうことなく、葉の老化時に野生型の濃度よりも高いトレオニンを産生した。
【0081】
実施例1:葉の中のトレオニン濃度に関する試験
緑色の葉または黄変した葉に対してトレオニン濃度に関する試験を行なった。葉片を採取し、分析に使用し、測定は、葉片当たりのトレオニンの量に基づくか(すなわち、thrの量/葉の面積)、上澄み中のタンパク質の量と関連づけられた(すなわち、トレオニンの量/1mgのタンパク質)。葉片を規定の体積の水と一緒にすり潰し、遠心分離して不溶な葉のくずを沈降させた。次いで、このプロセスで得られた上澄みをPhenomenex EZfaast Kitを用いて処理した。これは、標準的なLC/MS装置を用いて定量化することができるように抽出物中のアミノ酸を誘導体化するために使用される専売キットである。
【0082】
較正は、各アミノ酸の定量に関しては外部標準によって行い、誘導体化工程の効率は、内部標準のプロセスに含めることによって試料間で正規化した。クロマトグラムは、ピーク面積で評価し、LC/MSソフトウェアのアルゴリズムにおける積分を用いて濃度と関連づけた。ソフトウェアまたは操作者がピークの特定に確信が持てない場合には、「検出限界未満」と記載される。これは、空のベクター対照試料のいくつかと次世代で分離する無効な(null)植物のいくつかの場合であることが分かった。
【0083】
実施例2:葉特異的プロモーターに作動可能に連結されたフィードバック非感受性AK:HSDHによる形質転換植物
本発明者らは、シロイヌナズナ(At4g19710)由来の二機能性AK:HSDH野生型配列に対して部位特異的突然変異誘発を行なった。従って、最初に下記プライマーを用いるPCRによってシロイヌナズナ由来の葉特異的cDNAライブラリーから野生型配列を単離した:
正方向At4g19710 ATGGCGACTCTGAAGCCGTCATTTAC(配列番号8);
逆方向At4g19710 TTAAGACGGTGCACCGAGATAAGATGC(配列番号9)。
【0084】
この野生型配列を、以下の3つの方法のいずれか1つによって修飾した:(i)AK領域のみを突然変異させるか、(ii)HSDH領域を突然させるか、あるいは(iii)両方の領域を突然変異させて、Thr結合による調節を不可能にした。特異的突然変異は、どちらも酵素調節領域にあるGln443およびGln524に対して行ない、その両方を、アラニンに対する部位特異的突然変異誘発によって突然変異させた(Paris et al (2003), “Mechanism of control of Arabidopsis thaliana aspartate kinase-homoserine dehydrogenase by threonine”, J.Biol.Chem 278:5361-5366およびFrankard et al (1992), “Two feedback-insensitive enzymes of the aspartate pathway in Nicotiana sylvestris” Plant Physiol 99:1285-1293)。
【0085】
この手順のために、Stratagene(登録商標)社製QuikChange(登録商標)部位特異的変異誘発キット(Site-Directed Mutagenesis Kit)(カタログ番号200518)を使用した。部位特異的突然変異誘発反応において、Glu443に対するコドンコードを変えるために、下記プライマーを使用した:
正方向Gln443 GTGATTATGATATCAGCTGCTAGCAGTGAGCATTCTG(配列番号10);および
逆方向Gln443 CAGAATGCTCACTGCTAGCAGCTGATATCATAATTCAC(配列番号11)。
【0086】
Gln524に対して、下記プライマー対を使用した:
正方向Gln524 GTCCGAGCTATATCTGCTGGTTGTTCTGAGTACAATG(配列番号12);および
逆方向Gln524 CATTGATCTCAGAACAACCAGCAGATATAGCTCGGAC(配列番号13)。
【0087】
野生型シロイヌナズナAK:HSDHと同様に、これらの3つの突然変異型配列を個別に使用して、タバコ(Nicotiana tabacum)植物を形質転換した。全ての場合に、目的とする遺伝子は、葉特異的エンドウマメ・プラストシアニンプロモーターの制御下で発現された。アグロバクテリウム媒介形質転換を経て全ての個体群の植物を生成し、英国ケンブリッジにおいて温室条件下で育てた。EZfaastアミノ酸キット(Phenomenex(登録商標)社製)を使用して、各試料中の遊離アミノ酸を抽出および誘導体化した。次いで、LC/MSによって定量した。
【0088】
その結果を図5および表1に示す。図5では、AK領域のみが突然変異された一連の植物はAKで始まる名前を有し、HSDH領域のみが突然変異された一連の植物はHSDHで始まる名前を有し、AKおよびHSDH領域の両方が突然変異されている一連の植物はAK/HSDHで始まる名前を有し、野生型植物はWTで始まる名前を有し、空のベクター対照試料を含む形質転換植物はEVで始まる名前を有する。
【0089】
本発明者らは、非突然変異型シロイヌナズナ配列で形質転換された個体群を含む、シロイヌナズナ配列で形質転換された全ての個体群において葉のトレオニン濃度の上昇を達成した。AK領域で突然変異された配列で形質転換された個体群が葉の最も高いThr濃度を示した。これらは、適応度がひどく損なわれたことを示した最も高い割合の個体群と相関していた。本発明者らは、修飾による受精能への影響を抑えるつもりで葉特異的プロモーターを使用したが、それらが受けた影響は、植物全体が酵素に対するフィードバック制御の解除という代謝結果によってもなお影響を受けるには十分であった。
【0090】
しかし、トレオニンの上昇を示した全ての植物において、生育習性の変化との相関性があった。葉は、色が淡く、肥厚し、脆くかつ帯状であった。節間部は短く、成熟度が増すにつれて褐変を示した。芽は発育不良または奇形のいずれかであった。結論として、葉のトレオニンの上昇が得られた点では、部位特異的突然変異誘発は成功であった。しかし、葉に高濃度のトレオニンを有する農学的に生育可能な植物をこの手法から得るつもりであれば、アスパラギン酸キナーゼに対するフィードバック制御の解除による適応度コストをなお克服しなければならない。
【0091】
【表1】
実施例3:老化特異的プロモーターに作動可能に連結されたトレオニン非感受性AK:HSDHを含む形質転換植物
タバコ(Nicotiana tabacum)植物(品種K326)を使用して葉片を得、これを老化特異的プロモーターSAG12(その配列は図6、すなわち配列番号1に示されている)の制御下で、目的の遺伝子(すなわち、突然変異型AK:HSDH、図7(すなわち配列番号2、3および4)に示されているAK:HSDH配列)を保持するバイナリーベクターで(電気穿孔法によって)予め形質転換されていたアグロバクテリウム・ツメファシエンスと共培養した。バイナリーベクターがプロモーターを含むがAK:HSDH遺伝子を含まない対照個体群を同時に育てた。次いで、これらの葉片を組織培養プロトコルによって処理して小植物(Horsch et al. Science 227: 1229-1231, 1985)を得た。各小植物は、細菌からDNAを抽出し、植物のゲノムDNAに組み込む単一の形質転換イベントから得られたものであった。これらの植物を温室に移し、成熟するまで育てた。成熟した葉を植物から切り離し、ポリテン袋に入れて35℃で72時暗所に置いた。この時間の後、この葉を使用して葉片を得、上記実施例1に記載したように、トレオニン含有量を評価した。
【0092】
その結果を図8に示す。図8は、いくつかの植物において高濃度のトレオニンが得られたことを示している。有利なことに、これらの植物は正常に成長した。従って、これらの形質転換植物は、実施例2に記載されている植物で観察されたような適応度コストを示さなかった。
【0093】
実施例4:SAG12プロモーターに連結されたトレオニン非感受性AK:HSDHを含む形質転換植物の現場試験
SAG12:アスパラギン酸キナーゼ(pBNP 138-0253-001)の実験個体群から選択された植物細胞株をノースカロライナで2008年の間現場で育てた。この葉を火力乾燥し、選択した遊離アミノ酸の存在を分析した。この分析は、内部および外部標準較正によって確認されているLC/MSによるものであった。分析物が較正に使用される範囲を超える場合は、表2では「>S」として表中に示し、図では、関連する棒の上に星で示す。
【0094】
この試料一覧を「AK」番号に分類する。この番号は、本発明の遺伝子構築物で修飾されたK326バックグラウンド株、および3種類の代表的な対照試料、すなわち未修飾のK326、未修飾のKV1、未修飾のNC71であり、これら全てを同じ条件下、同じ時間で成長させ、かつ乾燥した。
【0095】
【表2】
図9を参照すると、3種類の対照試料(K326、KV1およびNC7)と比較した5種類の試験植物(AK1〜AK5)によって産生されたトレオニン濃度を示す棒グラフが示されている。図から分かるように、5種類の試験植物全てが、いずれの対照試料よりも著しく多いトレオニンを産生した。さらに、5種類の試験植物はいずれも、それらの成長適応度に対するコストを全く受けなかった。これらのデータは、本発明に係る構築物で形質転換された試験植物が商業用現場条件下で遊離トレオニンが(少なくとも2〜3倍)上昇した葉を生成し、これが乾燥プロセスの間も持続しているという観察を強く支持するものである。さらに、試験植物は、これらの現場条件下において全く不利な点を示さない非分離型であった。
【0096】
また、本発明者らは、現場試験植物から生成された乾葉中のいくつかの他のアミノ酸(GLN:グルタミン;GLU:グルタミン酸;ASN:アスパラギン;ASP:アスパラギン酸;HIS:ヒスチジン)の濃度も評価した。そのデータは図10〜14に示されている。
【0097】
図10から分かるように、細胞株AK3のグルタミン濃度は、3種類の対照細胞株の濃度に匹敵していた。図11に示すように、AK1〜3のグルタミン酸の量は対照試料と同じであったが、AK4〜5の濃度は僅かに高かった。図12は、アスパラギンの各濃度を示す。
【0098】
図13を参照すると、試験細胞株AK1〜AK3のアスパラギン酸濃度は、対照試料とほぼ同じであることが分かるが、AK4〜5の濃度は対照試料よりも高いように思われた。最後に図14を参照すると、ヒスチジン濃度は、試験細胞株AK4が濃度の上昇を示したこと以外は、全体においてほぼ同じであった。
【0099】
要約すると、図10〜14は、これらのアミノ酸の各濃度は対照試料に匹敵するか、対照試料よりもさらに高いことを明らかに示しており、これは、形質転換植物の適応度が損なわれなかったという良い兆候を示している。
【特許請求の範囲】
【請求項1】
トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物。
【請求項2】
前記老化特異的プロモーターが、シロイヌナズナの老化関連遺伝子から単離されたものである、請求項1に記載の遺伝子構築物。
【請求項3】
前記老化特異的プロモーターが、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは断片から選択される、請求項1または2に記載の遺伝子構築物。
【請求項4】
前記老化特異的プロモーターが、SAG12またはその機能的な変異体もしくは断片である、請求項3に記載の遺伝子構築物。
【請求項5】
前記プロモーターが、実質的に配列番号1に示されているようなヌクレオチド配列またはその機能的な変異体もしくは断片を含む、請求項3または4に記載の遺伝子構築物。
【請求項6】
前記ポリペプチドが、トレオニン非感受性アスパラギン酸キナーゼ(AK)またはその機能的な変異体もしくは断片である、前記請求項のいずれかに記載の遺伝子構築物。
【請求項7】
前記ポリペプチドが、トレオニン非感受性二機能性アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)である、前記請求項に記載の遺伝子構築物。
【請求項8】
トレオニン非感受性アスパラギン酸キナーゼ活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来する、前記請求項のいずれかに記載の遺伝子構築物。
【請求項9】
トレオニン非感受性アスパラギン酸キナーゼ活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、好ましくはシロイヌナズナ(A.thaliana)に由来する、前記請求項のいずれかに記載の遺伝子構築物。
【請求項10】
前記AK−HSDHの少なくとも1つのトレオニン結合部位が突然変異されている、請求項7〜9のいずれか1項に記載の遺伝子構築物。
【請求項11】
前記シロイヌナズナAK−HSDHがGln443および/またはGln524で突然変異されている、請求項7〜10のいずれか1項に記載の遺伝子構築物。
【請求項12】
前記AK−HSDHがGln443およびGln524で突然変異されている、請求項7〜11のいずれか1項に記載の遺伝子構築物。
【請求項13】
請求項1〜12のいずれか1項に記載の遺伝子構築物を含むベクター。
【請求項14】
同じ条件下で栽培された対応する野生型植物よりも高濃度のトレオニンをその葉に蓄積させる形質転換植物の産生方法であって、
i)植物細胞を請求項1〜12のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターで形質転換することと、
ii)前記形質転換された細胞から植物を再生することと、
を含む方法。
【請求項15】
請求項1〜13のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターを含む細胞。
【請求項16】
請求項1〜9のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターを含む形質転換植物。
【請求項17】
前記植物がアブラナ科、イネ科またはナス科由来である、請求項16に記載の形質転換植物。
【請求項18】
前記植物がタバコ植物である、請求項16または17に記載の形質転換植物。
【請求項19】
請求項16〜18のいずれか1項に記載の形質転換植物から得ることができる植物繁殖用製品。
【請求項20】
前記植物繁殖用製品が種子である、請求項19に記載の植物繁殖用製品。
【請求項21】
同じ条件下で栽培された野生型植物から採取した収穫された葉の中の対応するトレオニン濃度よりも高濃度のトレオニンを含有する収穫された葉であって、前記葉が、請求項16〜18のいずれか1項に記載の形質転換植物から収穫されたものまたは請求項14に記載の方法によって生産されたものである葉。
【請求項22】
突然変異型タバコ植物から得られたトレオニン豊富なタバコを含む喫煙物品であって、前記突然変異体が老化した葉の中にトレオニンを過剰産出することができる喫煙物品。
【請求項23】
前記突然変異型タバコ植物が、請求項1〜12のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターで形質転換されている、請求項22に記載の喫煙物品。
【請求項24】
植物の適応度を損なうことなく対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、植物代謝を変化させて葉の老化開始後にトレオニンの産生の上昇を達成することを含む方法。
【請求項25】
前記方法が、前記植物を請求項1〜12のいずれか1項に記載の構築物または請求項13に記載のベクターで形質転換することを含む、請求項24に記載の方法。
【請求項1】
トレオニン非感受性アスパラギン酸キナーゼ活性を有するポリペプチドをコードするコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物。
【請求項2】
前記老化特異的プロモーターが、シロイヌナズナの老化関連遺伝子から単離されたものである、請求項1に記載の遺伝子構築物。
【請求項3】
前記老化特異的プロモーターが、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは断片から選択される、請求項1または2に記載の遺伝子構築物。
【請求項4】
前記老化特異的プロモーターが、SAG12またはその機能的な変異体もしくは断片である、請求項3に記載の遺伝子構築物。
【請求項5】
前記プロモーターが、実質的に配列番号1に示されているようなヌクレオチド配列またはその機能的な変異体もしくは断片を含む、請求項3または4に記載の遺伝子構築物。
【請求項6】
前記ポリペプチドが、トレオニン非感受性アスパラギン酸キナーゼ(AK)またはその機能的な変異体もしくは断片である、前記請求項のいずれかに記載の遺伝子構築物。
【請求項7】
前記ポリペプチドが、トレオニン非感受性二機能性アスパラギン酸キナーゼ−ホモセリン脱水素酵素(AK−HSDH)である、前記請求項に記載の遺伝子構築物。
【請求項8】
トレオニン非感受性アスパラギン酸キナーゼ活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来する、前記請求項のいずれかに記載の遺伝子構築物。
【請求項9】
トレオニン非感受性アスパラギン酸キナーゼ活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、好ましくはシロイヌナズナ(A.thaliana)に由来する、前記請求項のいずれかに記載の遺伝子構築物。
【請求項10】
前記AK−HSDHの少なくとも1つのトレオニン結合部位が突然変異されている、請求項7〜9のいずれか1項に記載の遺伝子構築物。
【請求項11】
前記シロイヌナズナAK−HSDHがGln443および/またはGln524で突然変異されている、請求項7〜10のいずれか1項に記載の遺伝子構築物。
【請求項12】
前記AK−HSDHがGln443およびGln524で突然変異されている、請求項7〜11のいずれか1項に記載の遺伝子構築物。
【請求項13】
請求項1〜12のいずれか1項に記載の遺伝子構築物を含むベクター。
【請求項14】
同じ条件下で栽培された対応する野生型植物よりも高濃度のトレオニンをその葉に蓄積させる形質転換植物の産生方法であって、
i)植物細胞を請求項1〜12のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターで形質転換することと、
ii)前記形質転換された細胞から植物を再生することと、
を含む方法。
【請求項15】
請求項1〜13のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターを含む細胞。
【請求項16】
請求項1〜9のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターを含む形質転換植物。
【請求項17】
前記植物がアブラナ科、イネ科またはナス科由来である、請求項16に記載の形質転換植物。
【請求項18】
前記植物がタバコ植物である、請求項16または17に記載の形質転換植物。
【請求項19】
請求項16〜18のいずれか1項に記載の形質転換植物から得ることができる植物繁殖用製品。
【請求項20】
前記植物繁殖用製品が種子である、請求項19に記載の植物繁殖用製品。
【請求項21】
同じ条件下で栽培された野生型植物から採取した収穫された葉の中の対応するトレオニン濃度よりも高濃度のトレオニンを含有する収穫された葉であって、前記葉が、請求項16〜18のいずれか1項に記載の形質転換植物から収穫されたものまたは請求項14に記載の方法によって生産されたものである葉。
【請求項22】
突然変異型タバコ植物から得られたトレオニン豊富なタバコを含む喫煙物品であって、前記突然変異体が老化した葉の中にトレオニンを過剰産出することができる喫煙物品。
【請求項23】
前記突然変異型タバコ植物が、請求項1〜12のいずれか1項に記載の遺伝子構築物または請求項13に記載のベクターで形質転換されている、請求項22に記載の喫煙物品。
【請求項24】
植物の適応度を損なうことなく対応する野生型の濃度よりも植物の葉の中のトレオニン濃度を上昇させる方法であって、植物代謝を変化させて葉の老化開始後にトレオニンの産生の上昇を達成することを含む方法。
【請求項25】
前記方法が、前記植物を請求項1〜12のいずれか1項に記載の構築物または請求項13に記載のベクターで形質転換することを含む、請求項24に記載の方法。
【図1】

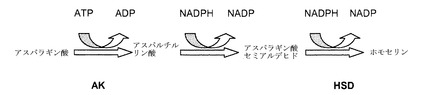
【図2】
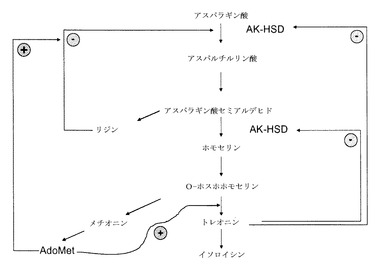
【図3】
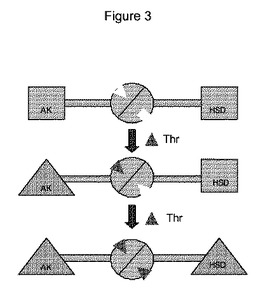
【図4】
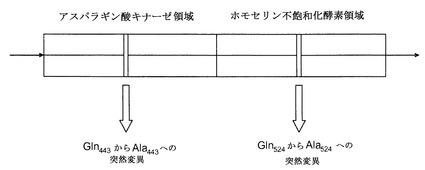
【図5】
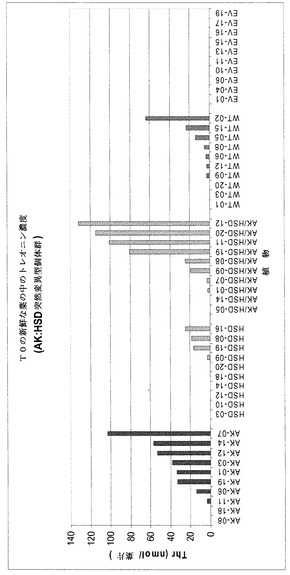
【図6】

【図7a】

【図7b】

【図8】
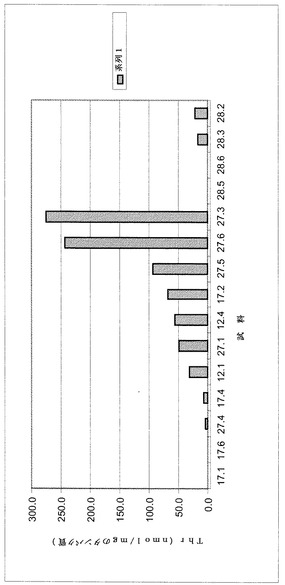
【図9】
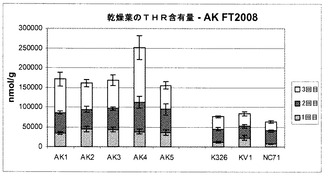
【図10】
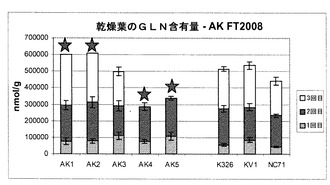
【図11】
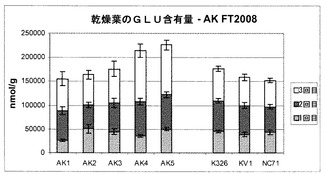
【図12】
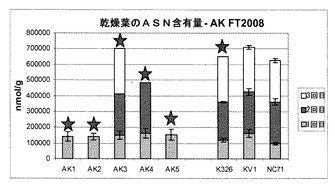
【図13】
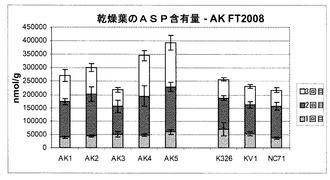
【図14】
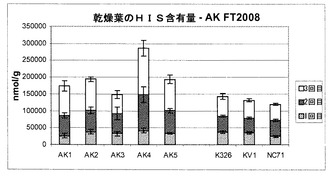
【図15】
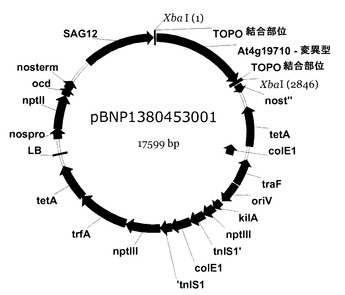
【図16】
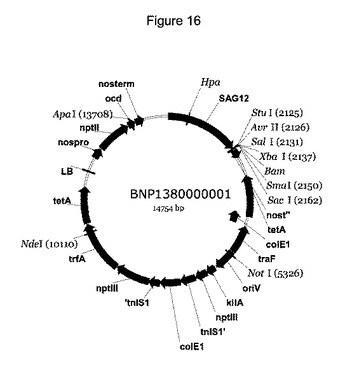
【図2】
【図3】
【図4】
【図5】
【図6】
【図7a】
【図7b】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2012−500003(P2012−500003A)
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願番号】特願2011−522524(P2011−522524)
【出願日】平成21年8月14日(2009.8.14)
【国際出願番号】PCT/EP2009/060582
【国際公開番号】WO2010/018234
【国際公開日】平成22年2月18日(2010.2.18)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願日】平成21年8月14日(2009.8.14)
【国際出願番号】PCT/EP2009/060582
【国際公開番号】WO2010/018234
【国際公開日】平成22年2月18日(2010.2.18)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
[ Back to top ]