
抗インフルエンザウイルス剤及びその製造方法
【課題】単独の植物から得られた成分であって、なおかつ高いインフルエンザウイルスに対する不活化効果を有する成分を含む抗インフルエンザ剤及びその製造方法を提供する。
【解決手段】ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤。抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法。前記植物果実の粉砕物を水中で煮沸し、水抽出液を固形分から分離することを含む。
【解決手段】ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤。抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法。前記植物果実の粉砕物を水中で煮沸し、水抽出液を固形分から分離することを含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗インフルエンザウイルス剤及びその製造方法に関する。
【背景技術】
【0002】
鳥インフルエンザ( Avian Influenza : AI )は、アジアを中心とする世界各国で発生している家禽の重要な感染症である。近年では人への感染・死亡例も多数報告されており、1997 年の香港における最初の発生ではH5N1 亜型ウイルスが18 人に重症呼吸器疾患を引き起こし、死亡者も出た。2003年から始まった東アジア一帯でのH5N1亜型の流行では多数の感染者、死亡者が報告されている。また、日本においても複数の地域の養鶏場で発生が確認された。このようにAI は、養鶏産業における経済的損失を引き起こすばかりでなく、人への感染の危険性も伴うことから発生拡大が強く懸念されている 。
【0003】
こうした脅威に備え、プレパンデミックワクチンが用意されてはいるものの、量が不足している上に新型ウイルスに対する効果は不透明である。新型ウイルスに適合したワクチンが開発・供給できるようになるまでには、パンデミック発生から数ヶ月を要する。一方、オセルタミビル(タミフル)やザナミビル(リレンザ)などのウイルス増殖を抑える抗ウイルス薬も実用化されているが、その有効性や耐性株の出現などの観点からの問題点も多い。このような背景から、近年新たな抗ウイルス薬の開発を目指し、さまざまな物質の抗ウイルス効果が検討されている。特に植物は古来より様々な薬効成分が発見され利用されてきた。天然物である植物は利用するにあたって安全性も高く、その成分は抗ウイルス薬として有効に利用できると期待されている。現在抗ウイルス薬として利用されているオセルタミビルも、かつて香辛料の八角から発見された。
【0004】
植物由来天然物の研究の結果、抗インフルエンザ作用を示す新しい植物由来物質として、これまでに複数の植物が報告されてきた。例えば、特許文献1にはアガリクス茸の親水性溶媒抽出物とスグリ属またはスノキ属植物の成分とを併用する抗インフルエンザ剤が記載されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2011-168572号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかし、特許文献1に記載の抗インフルエンザ剤が有する抗インフルエンザウイルス効果はさほど強いものではない。また、特許文献1に記載の抗インフルエンザ剤においては、アガリクス茸の親水性溶媒抽出物が主成分であり、アガリクス茸以外の植物成分を単独で抗インフルエンザ剤として使用できることも記載されていない。
【0007】
本発明は、単独の植物から得られた成分であって、なおかつ高いインフルエンザウイルスに対する不活化効果を有する成分を含む抗インフルエンザ剤を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らが種々検討した結果、ハスカップ、コケモモまたはマタタビの水抽出物が、H5N1亜型鳥インフルエンザウイルスに対して、極めて高い抗鳥インフルエンザウイルス活性を有することを見出して本発明を完成させた。
【0009】
本発明は、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤に関する。さらに本発明は、抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法であって、前記植物果実の粉砕または裁断物を水中で煮沸し、水抽出液を固形分から分離することを含む、前記製造方法に関する。
【発明の効果】
【0010】
本発明によれば、単独の植物から得られた成分であって、なおかつ高いインフルエンザウイルスに対する不活化効果を有する成分を含む抗インフルエンザ剤を提供することができる。
【図面の簡単な説明】
【0011】
【図1】エキス処理によるH5N1(山口株)ウイルス粒子の電子顕微鏡写真を示す。
【図2】蛍光抗体法による細胞内のウイルス蛋白の検出結果を示す。
【図3】Real-time RT-PCRによる細胞内のウイルス遺伝子の検出結果を示す。
【図4】エキスによるMDCK細胞への細胞障害性の検討(LDH assay) 結果を示す。
【発明を実施するための形態】
【0012】
<抗インフルエンザウイルス剤>
本発明の抗インフルエンザウイルス剤は、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する。
【0013】
ハスカップは、スイカズラ科スイカズラ属の落葉低木であり、その実(果実)は食用されている。本発明では、ハスカップの実(果実)を水抽出に用いる。水抽出に用いる実(果実)は、ハスカップの実の冷凍品またはその解凍品であっても良い。
コケモモは、ツツジ科スノキ属の常緑小低木であり、栽培されることは多くない。普通は,野性の実を採取し,その実(果実)は食用されている。本発明では、コケモモの果実を水抽出に用いる。
マタタビは、マタタビ科マタタビ属の落葉蔓性木本である。マタタビの実(果実)は薬用酒(マタタビ酒)の素として使われることがある。
ハスカップ、コケモモ及びマタタビの果実は、いずれも栽培品であっても、野生の果実であってもよい。
【0014】
これらの群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法は後述する。前記水抽出物は、例えば、前記植物果実の粉砕または裁断物の水抽出物であることができる。さらに、前記水抽出物は、熱水抽出物であることができる。本発明の抗インフルエンザウイルス剤は、前記水抽出物に対する希釈剤をさらに含有することもできる。希釈剤は特に制限はないが、例えば、水であることができる。
【0015】
本発明の抗インフルエンザウイルス剤は、インフルエンザウイルスに対する感染を予防し、またはインフルエンザウイルスに感染したヒトを含む動物(例えば、哺乳類、鳥類等)に対する治療するために用いることかできる。従って本発明の抗インフルエンザウイルス剤は、例えば、インフルエンザ治療薬、インフルエンザ感染予防薬、インフルエンザ感染予防用組成物として用いることができる。インフルエンザ感染予防用組成物は、例えば、手等の身体の消毒用や、インフルエンザウイルスの存在が疑われる物品の消毒、例えば、生活用品や調度品等の表面の消毒用として用いることができる。対象とするインフルエンザウイルスには特に制限はないが、例えば、鳥インフルエンザウイルスであることができる。
【0016】
本発明の抗インフルエンザウイルス剤をインフルエンザ治療薬またはインフルエンザ感染予防薬として用いる場合、本発明の抗インフルエンザウイルス剤の有効成分である水抽出物を、医師等の診断に基づいて、体重1kg当たり、例えば、0.1〜1,000mg、1日1回または2〜3回に分けて経口投与することができる。有効成分である水抽出物は、粉末または錠剤とすることができる他、水溶液とすることもできる。また投与形態は、経口に限らず、注射等も利用できる。
【0017】
本発明の抗インフルエンザウイルス剤をインフルエンザ感染予防用組成物として用いる場合、本発明の抗インフルエンザウイルス剤の有効成分である水抽出物を含有する水溶液等の液体であることができる。液体状のインフルエンザ感染予防用組成物を、例えば、手やインフルエンザウイルスの存在が疑われる物品に噴霧等することで、インフルエンザウイルスを死滅させることができる。インフルエンザ感染予防用組成物中の有効成分である水抽出物の濃度は、水抽出物に用いた果実の種類等を考慮して適宜決定することができる。
【0018】
<製造方法>
本発明の抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物は、例えば、前記植物果実を水中で煮沸し、水抽出液を固形分から分離することを含む方法で製造することができる。抽出効率という観点からは、植物果実は、粉砕または裁断等した物であることが好ましい。粉砕または裁断等した物の寸法特に制限はなく、植物の種類にもよるが、例えば、最も長い部分の長さを5 mm以内とすることができる。また、植物果実は、水抽出前に適宜乾燥することもできる。
【0019】
植物果実からの抽出は、常温の水中でも可能であるが、抽出完了までに長時間を要することから、熱水、例えば、50〜沸騰温度(例えば、100℃)の範囲の熱水で行うことが、比較的短時間で有効成分を抽出できるという観点から好ましい。より好ましくは、沸騰水での抽出である。さらに、熱水は、抽出と同時に水分を一部蒸発させて、抽出成分が濃縮されることから、有効成分が高い濃度で含有される抽出液が得られるという観点から好ましい。水分の蒸発の程度は、特に制限はないが、抽出開始時に使用した水の10〜90%の範囲の水を蒸発させることができ、好ましくは30〜70%の範囲の水を蒸発させることである。但し、有効成分の濃度によってウイルス不活性化の強弱があることから、不活性化すべきウイルスの種類や不活性化の程度、さらには、水を蒸発させて得られた抽出物の希釈の程度を考慮して、水の蒸発による濃縮の程度は適宜決定することができる。
【0020】
抽出終了後は、水抽出液を固形分から分離する。この分離のための方法は、特に制限はなく、例えば、ろ過及び遠心分離の少なくとも一方またはこれらを組み合わせて用いることができる。
【0021】
前記水抽出液は、乾燥して水抽出物の乾燥品を得ることもできる。乾燥方法には特に制限はないが、有効成分の熱による劣化等を防止しつつ乾燥品を得るという観点からは、フリーズドライ等の方法を用いる事が好ましい。
【実施例】
【0022】
以下本発明を実施例によりさらに詳細に説明する。但し、本発明は以下の実施例に限定されるものではない。以下の実施例ではハスカップ、コケモモおよびマタタビの各果実抽出液による抗H5N1亜型鳥インフルエンザウイルス効果について検討した。
【0023】
<実験方法>
1. 果実抽出液(エキス)
ハスカップとマタタビは栽培品、コケモモは天然品を用いた。それぞれの果実を乾燥させ、一辺が5mm以下になるように裁断したものを超純水1Lに対して20gの割合で加え、1時間半煮沸し全量を1/2とした。ガーゼを通して残渣を除いて得られた液を20,000 rpmで1時間遠心し、その上清をさらに5μmのフィルター付き遠心チューブでろ過したものを各エキス原液とした。エキスの希釈には超純水を使用した。対照として超純水を使用した。以下の実験では、エキス原液またはエキスの希釈液を用いた
【0024】
2. ウイルス、細胞
本実験において検討対象としたウイルスは、鳥インフルエンザウイルスである[A/chicken/Yamaguchi/7/2004(H5N1亜型)](山口株)、[A/whooper swan/Hokkaido/1/2008(H5N1亜型)](北海道株) である。ウイルスの希釈にはWHOのマニュアルに則って作製したウイルス増殖培地(VGM)を使用した。全てのウイルスはMDCK細胞および10日齢発育鶏卵に感染させた。
【0025】
3. エキス処理ウイルスのウイルス力価の測定
ウイルス原液と各エキス希釈液を等量混合し、混合後直ちに室温で一定時間静置したのち、VGMで10倍段階希釈して96 well組織培養用マイクロプレートに培養したMDCK細胞に接種した。ウイルスを接種後3日間インキュベーター内で培養し、3日目に各wellから回収した培養液に等量の0.5%鶏赤血球液を混合させ、赤血球凝集(HA)作用の有無を確認した。感染の有無はHA性の有無により判定し、50%組織培養感染量(TCID50)をBehrens-Karber法によって算出し、ウイルス力価とした。
細胞ではなく10日齢発育鶏卵を使用した実験では、作製したウイルスとエキスの混合液を同様に10倍段階希釈し、それぞれ100μlずつ漿尿膜腔内に接種した。3日後に漿尿膜腔液を採取し、HA性の有無から同様に50%発育鶏卵感染量(EID50)を算出した。
【0026】
4. エキスの細胞障害性の検討(LDH assay)
各エキスのMDCK細胞に対する細胞障害性を調べた。MDCK細胞に各エキス原液、および各エキスをVGMで2倍、4倍、8倍希釈した液(エキス含培地)をそれぞれ添加し、3日間培養した。回収した培養上清中に含まれるLactate dehydrogenase (LDH) の量をCytotoxicity Detection kit (Roche, Mannheim, Germany) を用いて測定した。得られた値を基に各エキスのMDCK細胞に対する細胞傷害性を算出した。
【0027】
5. ウイルス感染細胞におけるエキスによるウイルス増殖抑制効果の検討
H5N1(山口株)ウイルス液をVGMで10倍段階希釈した後、96 well組織培養用マイクロプレートに培養したMDCK細胞に接種し、37℃に1時間おいてウイルスを吸着させた。その後細胞表面をPBSで3回洗浄しウイルス液を除いた後、各エキスをVGMでそれぞれ2倍、5倍、10倍希釈した液(エキス含培地)を加えて3日間培養し、TCID50を同様に算出した。
【0028】
6. 赤血球凝集阻害試験
ウイルスの細胞吸着能がエキスにより消失するかどうかを検討するため、赤血球凝集(HA)試験を行った。各エキスの原液を96 well-V底プレート上でH5N1(山口株)ウイルス液と等量混合し、室温で10分間反応させた。反応後、その混合液をプレート上でVGMにて2倍段階希釈した。その後、0.5%鶏赤血球液を全てのwellに等量添加し、室温で1時間静置後,赤血球凝集反応を確認した。完全な赤血球凝集を引き起こすウイルスの最高希釈率を,HA 価として決定した。
【0029】
7. 蛍光抗体法によるウイルス蛋白の検出
5. により得られた細胞中のウイルス蛋白を蛍光抗体法により検出した。エキス含有培地には,各エキスの5倍希釈したものを使用した。細胞をアセトンで固定したのち、一次抗体としてマウス抗A型インフルエンザNP抗体を添加し、37℃で30分反応させた。その後PBSで洗浄し、二次抗体として蛍光標識ウサギ抗マウスIgGを添加し、37℃で30分反応させた。再びPBSで洗浄し50%グリセロールPBSを加えた後、励起波長492 nmにて蛍光顕微鏡で観察した。
【0030】
8. Real-time RT-PCRによるウイルス遺伝子の検出
5. により得られた細胞中のウイルス遺伝子をReal-time RT-PCRにより検出した。エキス含有培地はハスカップおよびコケモモエキスを5倍希釈したものを使用した。ウイルスRNAをMagMaxTM-96 AI/ND Viral RNA Isolation Kit (Ambion Inc. Austin, TX) 及びKingFisher Purification System(Thermo Scientific, Waltham, MA) を用いて分離し、random primer, oligo(dt)primer及びMMLV reverse transcriptase(Invitrogen. Carlsbad, CA)を用いてRNAから逆転写反応を行い、cDNAを作製した。逆転写反応は25℃ 10分、37℃ 50分、65℃ 10分の条件下で行った。得られたcDNA及びEagleTaq Master Mix kit(Roche, Branchburg, NJ)を用いてABI PRISM Sequence Detection System 7500HT(Applied Biosystems, Foster, CA)においてreal-time PCRを行った。
【0031】
M遺伝子検出のために、以下のプライマーを使用した。
Probe: 6-FAM-TCAGGCCCCCTCAAAGCCGA-TAMRA (配列番号1)
Forward: 5'-ARATGAGTCTTCTRACCGAGGTCG-3'(配列番号2)
Reverse: 5'-TGCAAAGACATCYTCAAGYYTCTG-3'(配列番号3)
【0032】
内在性コントロールとして,MDCK細胞由来のハウスキーピング遺伝子をEukaryotic 18S rRNA Endogenous Control (Applied Biosystems)を用いて検出した。
【0033】
Real-time PCRは95℃ 10分間でcDNAの変性を行った後、95℃ 15秒、60℃ 1分の反応を45回繰り返す条件下において行った。Real-time PCRで各サンプルの蛍光強度が一定値を超えたときのサイクル数を求めることにより、各サンプルの閾値サイクル数(Ct)を算出し、内在性コントロールの値との相対比を求めた。
【0034】
9. 電子顕微鏡によるウイルス粒子形態の解析
電子顕微鏡を用いて、各エキスで処理したH5N1(山口株)ウイルス粒子の形態学的変化を観察した。カーボン蒸着コロジオン膜マイクログリッド(日新EM, 東京, 日本)に、各エキス原液と等量混合することで処理したH5N1(山口株)ウイルス液を,室温でマイクログリッド上のコロジオン膜に5分間吸着させた。その後2% リンタングステン酸溶液(pH 6.5)によって2分間陰性染色し、H7500電子顕微鏡(日立,東京,日本)を用いて観察した。
【0035】
<実験結果>
1. 果実エキス処理によるH5N1ウイルス力価の低下
H5N1ウイルス(山口株・北海道株)と各果実エキスの原液を等量混合後、直ちに10日齢発育鶏卵に接種した場合(0分処理)のウイルス力価(log10 EID50/ml)を測定した。H5N1(山口株)ウイルスについては、全てのエキスによる処理によってウイルス力価は,検出限界(1.5log10)以下に低下した。H5N1(北海道株)ウイルスについては、ウイルス対照のウイルス力価と比較して、ハスカップエキスによる処理で5.0 log10、コケモモおよびマタタビエキスによる処理で4.5 log10ウイルス力価が低下した。
【0036】
【表1】

【0037】
各エキスはエキス原液による0分処理により、H5N1(山口株)ウイルスの力価を検出限界以下まで低下させたことから,非常に高い不活化効果を示した。一方,H5N1北海道株に対しては山口株と比較すると,各エキスの不活化効果が低かったが,ウイルス対照と比較して104.5から105.0倍ウイルス力価が低下したことから,十分な不活化効果を示していると思われる。
【0038】
2.赤血球凝集阻害試験
ウイルスの細胞吸着能がエキスにより消失するかどうかを検討するため、HA試験を行った。各エキス原液での10分間処理により、H5N1(山口株)ウイルスのHA価は,陽性限界以下(HA価8倍以下)に低下した。PBSと混合したウイルス対照ではHA価は64倍であった。
【0039】
【表2】
【0040】
H5N1(山口株)ウイルスを用いた赤血球凝集阻害試験では、各エキスにはH5N1ウイルスに対する高い赤血球凝集阻害作用がみられた。HA活性の低下は、ウイルス不活化効果と密接に関連していることが示唆された。HA活性を失ったウイルスは細胞に吸着できず感染が成立しないことからも、HAに対する阻害作用が、エキスによるウイルス不活化の主要な機構となっていると考えられた。
【0041】
3. 電子顕微鏡によるウイルス粒子の形態観察
エキス処理によるH5N1(山口株)ウイルス粒子の形態学的変化について電子顕微鏡を用いて観察した。電子顕微鏡写真を図1に示す。未処理H5N1ウイルスでは表面にスパイク蛋白が認められ、構造が比較的均一かつ明瞭に認められる楕円形のウイルス粒子が観察された。一方、ハスカップエキスで処理したサンプルでは,正常なウイルス粒子を見つけることが出来なかった。H5N1(山口株)ウイルスのウイルス粒子は、未処理H5N1ウイルスと比較して、粒子表面のスパイク蛋白が明瞭には認められず、粒子全体が不整形に変形しており、複数のウイルス粒子が凝集して存在しているように観察された。また,他のエキス処理サンプルにおいても,同様な変化が認められた。
【0042】
エキスで処理したH5N1(山口株)ウイルスのサンプルでは,正常な形態を持つウイルス粒子を見つけることが出来なかったことから,エキスの作用によるウイルス粒子の破壊が起きている可能性が示唆される。このこともウイルスの不活化を引き起こす,大きな要因となっていると思われる。
【0043】
4. ウイルス感染細胞におけるウイルス増殖抑制効果の検討
ウイルス感染細胞を各エキスを含む培地で培養することで、各エキスのウイルス増殖抑制効果を検討した。2倍希釈のエキスを含む培地(2倍エキス含培地)での培養により、全てのエキスでH5N1(山口株)ウイルスのウイルス力価が検出限界(1.5 log10)以下まで低下した。5倍エキス含培地での培養では、ハスカップでのみウイルス力価が検出限界(1.5 log10)以下まで低下したが、コケモモ、マタタビでは,有意なウイルス力価の低下はみられなかった。
【0044】
【表3】
【0045】
ウイルス感染後の細胞にエキスを処理した実験では、2倍(ハスカップのみ5倍)希釈までのエキスを含む培地での培養によってウイルスの増殖が完全に抑えられていたことから、各エキスにはウイルスと混合することでウイルスを直接不活化するだけではなく、細胞に感染して細胞内に入り込んだウイルスに対しても増殖を抑制する効果があることが確認できた。
【0046】
5. 蛍光抗体法による細胞内のウイルス蛋白の検出
4.においてウイルス増殖が抑えられていた細胞内でのウイルス蛋白合成について、蛍光抗体法により検証した。結果を図2に示す。4.においてウイルス増殖抑制効果がみられたハスカップの5倍希釈のエキスを含むエキス含培地により培養した細胞では、ウイルス蛋白が全く検出されなかった。一方,他のエキスの5倍希釈ではウイルス対照と同程度のウイルス蛋白が検出された。
【0047】
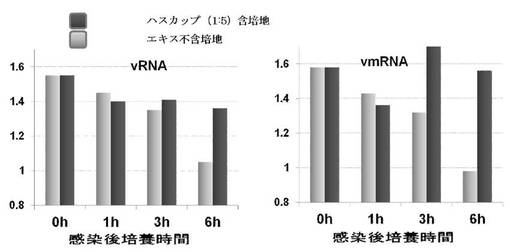
6. Real-time RT-PCRによる細胞内のウイルス遺伝子の検出
4.において,ハスカップおよびコケモモエキス含培地(5倍希釈)により培養した細胞内でのvRNAおよびvmRNA合成について、Real-time RT-PCRを用いて解析した。結果を図3に示す。得られたCt は、MDCK細胞由来のハウスキーピング遺伝子であるGAPDHのCtとの相対比で表すことにより補正した(グラフの縦軸)。エキス不含培地で培養した細胞では、vRNAおよびvmRNAについてのCtが培養0時間から24時間まで継時的に低下し、vRNAおよびvmRNA合成が進行していることが確認された。一方、ハスカップエキス 含培地で培養した細胞では、vRNA、vmRNAともに培養1時間まではエキス不含培地で培養した細胞と同程度のCtであったが、培養3時間および6時間では、エキス不含培地で培養した細胞と比べ高いCtであった。しかし、培養24時間ではエキス不含培地で培養した細胞と同程度までCtが低下した。
【0048】
コケモモエキス含培地で培養した細胞では、vRNA、vmRNAともに培養1時間まではエキス不含培地で培養した細胞と比べCtが大きく低下した。培養3時間から24時間まではエキス不含培地で培養した細胞と比べやや高い値を示しつつ低下した(データ示さず)。
【0049】
感染細胞内でのウイルス動態を知る目的で、蛍光抗体法及びreal-time RT-PCRを用いて解析を行った。蛍光抗体法の結果から、ハスカップエキス存在下での培養では、感染細胞中でウイルス蛋白の合成が進行していないことが示された。また,real-time RT-PCRによって、ハスカップエキス含培地で培養・維持された感染細胞中でウイルスRNAの転写の進行が抑制されていることが示された。しかし,24時間後では,ウイルス対照のCt値と差が認められなかったことから,ウイルスRNAの産生は抑制されているものの,完全には阻害されていないと思われる。一方,ウイルス蛋白が合成されていないことから,ウイルス遺伝子からの蛋白質の翻訳が何らかのメカニズムで阻害されていることが示唆された。
【0050】
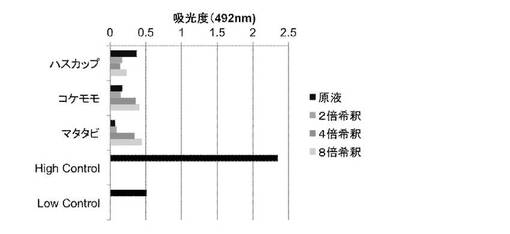
7. エキスによるMDCK細胞への細胞障害性の検討(LDH assay)
MDCK細胞への細胞障害性について、LDH assayにより検討した。結果を図4に示す。原液から8倍希釈まで、全てのエキスにおいて細胞障害性は認められなかった。
【0051】
果実エキスは、少なくとも原液から8倍濃度までの範囲の濃度では細胞障害性を全く示さなかった。さらに、5倍希釈のエキスにより2日間培養した細胞は、その後問題なく継代することができた(データ示さず)。これらのことから、果実エキスは、細胞に対して影響を与ることなく、細胞中のウイルスの複製に対してのみ影響を及ぼしていたことになる。
【産業上の利用可能性】
【0052】
本発明は、インフルエンザウイルスによる感染治療及び予防の分野に有用である。
【技術分野】
【0001】
本発明は、抗インフルエンザウイルス剤及びその製造方法に関する。
【背景技術】
【0002】
鳥インフルエンザ( Avian Influenza : AI )は、アジアを中心とする世界各国で発生している家禽の重要な感染症である。近年では人への感染・死亡例も多数報告されており、1997 年の香港における最初の発生ではH5N1 亜型ウイルスが18 人に重症呼吸器疾患を引き起こし、死亡者も出た。2003年から始まった東アジア一帯でのH5N1亜型の流行では多数の感染者、死亡者が報告されている。また、日本においても複数の地域の養鶏場で発生が確認された。このようにAI は、養鶏産業における経済的損失を引き起こすばかりでなく、人への感染の危険性も伴うことから発生拡大が強く懸念されている 。
【0003】
こうした脅威に備え、プレパンデミックワクチンが用意されてはいるものの、量が不足している上に新型ウイルスに対する効果は不透明である。新型ウイルスに適合したワクチンが開発・供給できるようになるまでには、パンデミック発生から数ヶ月を要する。一方、オセルタミビル(タミフル)やザナミビル(リレンザ)などのウイルス増殖を抑える抗ウイルス薬も実用化されているが、その有効性や耐性株の出現などの観点からの問題点も多い。このような背景から、近年新たな抗ウイルス薬の開発を目指し、さまざまな物質の抗ウイルス効果が検討されている。特に植物は古来より様々な薬効成分が発見され利用されてきた。天然物である植物は利用するにあたって安全性も高く、その成分は抗ウイルス薬として有効に利用できると期待されている。現在抗ウイルス薬として利用されているオセルタミビルも、かつて香辛料の八角から発見された。
【0004】
植物由来天然物の研究の結果、抗インフルエンザ作用を示す新しい植物由来物質として、これまでに複数の植物が報告されてきた。例えば、特許文献1にはアガリクス茸の親水性溶媒抽出物とスグリ属またはスノキ属植物の成分とを併用する抗インフルエンザ剤が記載されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2011-168572号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかし、特許文献1に記載の抗インフルエンザ剤が有する抗インフルエンザウイルス効果はさほど強いものではない。また、特許文献1に記載の抗インフルエンザ剤においては、アガリクス茸の親水性溶媒抽出物が主成分であり、アガリクス茸以外の植物成分を単独で抗インフルエンザ剤として使用できることも記載されていない。
【0007】
本発明は、単独の植物から得られた成分であって、なおかつ高いインフルエンザウイルスに対する不活化効果を有する成分を含む抗インフルエンザ剤を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らが種々検討した結果、ハスカップ、コケモモまたはマタタビの水抽出物が、H5N1亜型鳥インフルエンザウイルスに対して、極めて高い抗鳥インフルエンザウイルス活性を有することを見出して本発明を完成させた。
【0009】
本発明は、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤に関する。さらに本発明は、抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法であって、前記植物果実の粉砕または裁断物を水中で煮沸し、水抽出液を固形分から分離することを含む、前記製造方法に関する。
【発明の効果】
【0010】
本発明によれば、単独の植物から得られた成分であって、なおかつ高いインフルエンザウイルスに対する不活化効果を有する成分を含む抗インフルエンザ剤を提供することができる。
【図面の簡単な説明】
【0011】
【図1】エキス処理によるH5N1(山口株)ウイルス粒子の電子顕微鏡写真を示す。
【図2】蛍光抗体法による細胞内のウイルス蛋白の検出結果を示す。
【図3】Real-time RT-PCRによる細胞内のウイルス遺伝子の検出結果を示す。
【図4】エキスによるMDCK細胞への細胞障害性の検討(LDH assay) 結果を示す。
【発明を実施するための形態】
【0012】
<抗インフルエンザウイルス剤>
本発明の抗インフルエンザウイルス剤は、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する。
【0013】
ハスカップは、スイカズラ科スイカズラ属の落葉低木であり、その実(果実)は食用されている。本発明では、ハスカップの実(果実)を水抽出に用いる。水抽出に用いる実(果実)は、ハスカップの実の冷凍品またはその解凍品であっても良い。
コケモモは、ツツジ科スノキ属の常緑小低木であり、栽培されることは多くない。普通は,野性の実を採取し,その実(果実)は食用されている。本発明では、コケモモの果実を水抽出に用いる。
マタタビは、マタタビ科マタタビ属の落葉蔓性木本である。マタタビの実(果実)は薬用酒(マタタビ酒)の素として使われることがある。
ハスカップ、コケモモ及びマタタビの果実は、いずれも栽培品であっても、野生の果実であってもよい。
【0014】
これらの群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法は後述する。前記水抽出物は、例えば、前記植物果実の粉砕または裁断物の水抽出物であることができる。さらに、前記水抽出物は、熱水抽出物であることができる。本発明の抗インフルエンザウイルス剤は、前記水抽出物に対する希釈剤をさらに含有することもできる。希釈剤は特に制限はないが、例えば、水であることができる。
【0015】
本発明の抗インフルエンザウイルス剤は、インフルエンザウイルスに対する感染を予防し、またはインフルエンザウイルスに感染したヒトを含む動物(例えば、哺乳類、鳥類等)に対する治療するために用いることかできる。従って本発明の抗インフルエンザウイルス剤は、例えば、インフルエンザ治療薬、インフルエンザ感染予防薬、インフルエンザ感染予防用組成物として用いることができる。インフルエンザ感染予防用組成物は、例えば、手等の身体の消毒用や、インフルエンザウイルスの存在が疑われる物品の消毒、例えば、生活用品や調度品等の表面の消毒用として用いることができる。対象とするインフルエンザウイルスには特に制限はないが、例えば、鳥インフルエンザウイルスであることができる。
【0016】
本発明の抗インフルエンザウイルス剤をインフルエンザ治療薬またはインフルエンザ感染予防薬として用いる場合、本発明の抗インフルエンザウイルス剤の有効成分である水抽出物を、医師等の診断に基づいて、体重1kg当たり、例えば、0.1〜1,000mg、1日1回または2〜3回に分けて経口投与することができる。有効成分である水抽出物は、粉末または錠剤とすることができる他、水溶液とすることもできる。また投与形態は、経口に限らず、注射等も利用できる。
【0017】
本発明の抗インフルエンザウイルス剤をインフルエンザ感染予防用組成物として用いる場合、本発明の抗インフルエンザウイルス剤の有効成分である水抽出物を含有する水溶液等の液体であることができる。液体状のインフルエンザ感染予防用組成物を、例えば、手やインフルエンザウイルスの存在が疑われる物品に噴霧等することで、インフルエンザウイルスを死滅させることができる。インフルエンザ感染予防用組成物中の有効成分である水抽出物の濃度は、水抽出物に用いた果実の種類等を考慮して適宜決定することができる。
【0018】
<製造方法>
本発明の抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物は、例えば、前記植物果実を水中で煮沸し、水抽出液を固形分から分離することを含む方法で製造することができる。抽出効率という観点からは、植物果実は、粉砕または裁断等した物であることが好ましい。粉砕または裁断等した物の寸法特に制限はなく、植物の種類にもよるが、例えば、最も長い部分の長さを5 mm以内とすることができる。また、植物果実は、水抽出前に適宜乾燥することもできる。
【0019】
植物果実からの抽出は、常温の水中でも可能であるが、抽出完了までに長時間を要することから、熱水、例えば、50〜沸騰温度(例えば、100℃)の範囲の熱水で行うことが、比較的短時間で有効成分を抽出できるという観点から好ましい。より好ましくは、沸騰水での抽出である。さらに、熱水は、抽出と同時に水分を一部蒸発させて、抽出成分が濃縮されることから、有効成分が高い濃度で含有される抽出液が得られるという観点から好ましい。水分の蒸発の程度は、特に制限はないが、抽出開始時に使用した水の10〜90%の範囲の水を蒸発させることができ、好ましくは30〜70%の範囲の水を蒸発させることである。但し、有効成分の濃度によってウイルス不活性化の強弱があることから、不活性化すべきウイルスの種類や不活性化の程度、さらには、水を蒸発させて得られた抽出物の希釈の程度を考慮して、水の蒸発による濃縮の程度は適宜決定することができる。
【0020】
抽出終了後は、水抽出液を固形分から分離する。この分離のための方法は、特に制限はなく、例えば、ろ過及び遠心分離の少なくとも一方またはこれらを組み合わせて用いることができる。
【0021】
前記水抽出液は、乾燥して水抽出物の乾燥品を得ることもできる。乾燥方法には特に制限はないが、有効成分の熱による劣化等を防止しつつ乾燥品を得るという観点からは、フリーズドライ等の方法を用いる事が好ましい。
【実施例】
【0022】
以下本発明を実施例によりさらに詳細に説明する。但し、本発明は以下の実施例に限定されるものではない。以下の実施例ではハスカップ、コケモモおよびマタタビの各果実抽出液による抗H5N1亜型鳥インフルエンザウイルス効果について検討した。
【0023】
<実験方法>
1. 果実抽出液(エキス)
ハスカップとマタタビは栽培品、コケモモは天然品を用いた。それぞれの果実を乾燥させ、一辺が5mm以下になるように裁断したものを超純水1Lに対して20gの割合で加え、1時間半煮沸し全量を1/2とした。ガーゼを通して残渣を除いて得られた液を20,000 rpmで1時間遠心し、その上清をさらに5μmのフィルター付き遠心チューブでろ過したものを各エキス原液とした。エキスの希釈には超純水を使用した。対照として超純水を使用した。以下の実験では、エキス原液またはエキスの希釈液を用いた
【0024】
2. ウイルス、細胞
本実験において検討対象としたウイルスは、鳥インフルエンザウイルスである[A/chicken/Yamaguchi/7/2004(H5N1亜型)](山口株)、[A/whooper swan/Hokkaido/1/2008(H5N1亜型)](北海道株) である。ウイルスの希釈にはWHOのマニュアルに則って作製したウイルス増殖培地(VGM)を使用した。全てのウイルスはMDCK細胞および10日齢発育鶏卵に感染させた。
【0025】
3. エキス処理ウイルスのウイルス力価の測定
ウイルス原液と各エキス希釈液を等量混合し、混合後直ちに室温で一定時間静置したのち、VGMで10倍段階希釈して96 well組織培養用マイクロプレートに培養したMDCK細胞に接種した。ウイルスを接種後3日間インキュベーター内で培養し、3日目に各wellから回収した培養液に等量の0.5%鶏赤血球液を混合させ、赤血球凝集(HA)作用の有無を確認した。感染の有無はHA性の有無により判定し、50%組織培養感染量(TCID50)をBehrens-Karber法によって算出し、ウイルス力価とした。
細胞ではなく10日齢発育鶏卵を使用した実験では、作製したウイルスとエキスの混合液を同様に10倍段階希釈し、それぞれ100μlずつ漿尿膜腔内に接種した。3日後に漿尿膜腔液を採取し、HA性の有無から同様に50%発育鶏卵感染量(EID50)を算出した。
【0026】
4. エキスの細胞障害性の検討(LDH assay)
各エキスのMDCK細胞に対する細胞障害性を調べた。MDCK細胞に各エキス原液、および各エキスをVGMで2倍、4倍、8倍希釈した液(エキス含培地)をそれぞれ添加し、3日間培養した。回収した培養上清中に含まれるLactate dehydrogenase (LDH) の量をCytotoxicity Detection kit (Roche, Mannheim, Germany) を用いて測定した。得られた値を基に各エキスのMDCK細胞に対する細胞傷害性を算出した。
【0027】
5. ウイルス感染細胞におけるエキスによるウイルス増殖抑制効果の検討
H5N1(山口株)ウイルス液をVGMで10倍段階希釈した後、96 well組織培養用マイクロプレートに培養したMDCK細胞に接種し、37℃に1時間おいてウイルスを吸着させた。その後細胞表面をPBSで3回洗浄しウイルス液を除いた後、各エキスをVGMでそれぞれ2倍、5倍、10倍希釈した液(エキス含培地)を加えて3日間培養し、TCID50を同様に算出した。
【0028】
6. 赤血球凝集阻害試験
ウイルスの細胞吸着能がエキスにより消失するかどうかを検討するため、赤血球凝集(HA)試験を行った。各エキスの原液を96 well-V底プレート上でH5N1(山口株)ウイルス液と等量混合し、室温で10分間反応させた。反応後、その混合液をプレート上でVGMにて2倍段階希釈した。その後、0.5%鶏赤血球液を全てのwellに等量添加し、室温で1時間静置後,赤血球凝集反応を確認した。完全な赤血球凝集を引き起こすウイルスの最高希釈率を,HA 価として決定した。
【0029】
7. 蛍光抗体法によるウイルス蛋白の検出
5. により得られた細胞中のウイルス蛋白を蛍光抗体法により検出した。エキス含有培地には,各エキスの5倍希釈したものを使用した。細胞をアセトンで固定したのち、一次抗体としてマウス抗A型インフルエンザNP抗体を添加し、37℃で30分反応させた。その後PBSで洗浄し、二次抗体として蛍光標識ウサギ抗マウスIgGを添加し、37℃で30分反応させた。再びPBSで洗浄し50%グリセロールPBSを加えた後、励起波長492 nmにて蛍光顕微鏡で観察した。
【0030】
8. Real-time RT-PCRによるウイルス遺伝子の検出
5. により得られた細胞中のウイルス遺伝子をReal-time RT-PCRにより検出した。エキス含有培地はハスカップおよびコケモモエキスを5倍希釈したものを使用した。ウイルスRNAをMagMaxTM-96 AI/ND Viral RNA Isolation Kit (Ambion Inc. Austin, TX) 及びKingFisher Purification System(Thermo Scientific, Waltham, MA) を用いて分離し、random primer, oligo(dt)primer及びMMLV reverse transcriptase(Invitrogen. Carlsbad, CA)を用いてRNAから逆転写反応を行い、cDNAを作製した。逆転写反応は25℃ 10分、37℃ 50分、65℃ 10分の条件下で行った。得られたcDNA及びEagleTaq Master Mix kit(Roche, Branchburg, NJ)を用いてABI PRISM Sequence Detection System 7500HT(Applied Biosystems, Foster, CA)においてreal-time PCRを行った。
【0031】
M遺伝子検出のために、以下のプライマーを使用した。
Probe: 6-FAM-TCAGGCCCCCTCAAAGCCGA-TAMRA (配列番号1)
Forward: 5'-ARATGAGTCTTCTRACCGAGGTCG-3'(配列番号2)
Reverse: 5'-TGCAAAGACATCYTCAAGYYTCTG-3'(配列番号3)
【0032】
内在性コントロールとして,MDCK細胞由来のハウスキーピング遺伝子をEukaryotic 18S rRNA Endogenous Control (Applied Biosystems)を用いて検出した。
【0033】
Real-time PCRは95℃ 10分間でcDNAの変性を行った後、95℃ 15秒、60℃ 1分の反応を45回繰り返す条件下において行った。Real-time PCRで各サンプルの蛍光強度が一定値を超えたときのサイクル数を求めることにより、各サンプルの閾値サイクル数(Ct)を算出し、内在性コントロールの値との相対比を求めた。
【0034】
9. 電子顕微鏡によるウイルス粒子形態の解析
電子顕微鏡を用いて、各エキスで処理したH5N1(山口株)ウイルス粒子の形態学的変化を観察した。カーボン蒸着コロジオン膜マイクログリッド(日新EM, 東京, 日本)に、各エキス原液と等量混合することで処理したH5N1(山口株)ウイルス液を,室温でマイクログリッド上のコロジオン膜に5分間吸着させた。その後2% リンタングステン酸溶液(pH 6.5)によって2分間陰性染色し、H7500電子顕微鏡(日立,東京,日本)を用いて観察した。
【0035】
<実験結果>
1. 果実エキス処理によるH5N1ウイルス力価の低下
H5N1ウイルス(山口株・北海道株)と各果実エキスの原液を等量混合後、直ちに10日齢発育鶏卵に接種した場合(0分処理)のウイルス力価(log10 EID50/ml)を測定した。H5N1(山口株)ウイルスについては、全てのエキスによる処理によってウイルス力価は,検出限界(1.5log10)以下に低下した。H5N1(北海道株)ウイルスについては、ウイルス対照のウイルス力価と比較して、ハスカップエキスによる処理で5.0 log10、コケモモおよびマタタビエキスによる処理で4.5 log10ウイルス力価が低下した。
【0036】
【表1】
【0037】
各エキスはエキス原液による0分処理により、H5N1(山口株)ウイルスの力価を検出限界以下まで低下させたことから,非常に高い不活化効果を示した。一方,H5N1北海道株に対しては山口株と比較すると,各エキスの不活化効果が低かったが,ウイルス対照と比較して104.5から105.0倍ウイルス力価が低下したことから,十分な不活化効果を示していると思われる。
【0038】
2.赤血球凝集阻害試験
ウイルスの細胞吸着能がエキスにより消失するかどうかを検討するため、HA試験を行った。各エキス原液での10分間処理により、H5N1(山口株)ウイルスのHA価は,陽性限界以下(HA価8倍以下)に低下した。PBSと混合したウイルス対照ではHA価は64倍であった。
【0039】
【表2】
【0040】
H5N1(山口株)ウイルスを用いた赤血球凝集阻害試験では、各エキスにはH5N1ウイルスに対する高い赤血球凝集阻害作用がみられた。HA活性の低下は、ウイルス不活化効果と密接に関連していることが示唆された。HA活性を失ったウイルスは細胞に吸着できず感染が成立しないことからも、HAに対する阻害作用が、エキスによるウイルス不活化の主要な機構となっていると考えられた。
【0041】
3. 電子顕微鏡によるウイルス粒子の形態観察
エキス処理によるH5N1(山口株)ウイルス粒子の形態学的変化について電子顕微鏡を用いて観察した。電子顕微鏡写真を図1に示す。未処理H5N1ウイルスでは表面にスパイク蛋白が認められ、構造が比較的均一かつ明瞭に認められる楕円形のウイルス粒子が観察された。一方、ハスカップエキスで処理したサンプルでは,正常なウイルス粒子を見つけることが出来なかった。H5N1(山口株)ウイルスのウイルス粒子は、未処理H5N1ウイルスと比較して、粒子表面のスパイク蛋白が明瞭には認められず、粒子全体が不整形に変形しており、複数のウイルス粒子が凝集して存在しているように観察された。また,他のエキス処理サンプルにおいても,同様な変化が認められた。
【0042】
エキスで処理したH5N1(山口株)ウイルスのサンプルでは,正常な形態を持つウイルス粒子を見つけることが出来なかったことから,エキスの作用によるウイルス粒子の破壊が起きている可能性が示唆される。このこともウイルスの不活化を引き起こす,大きな要因となっていると思われる。
【0043】
4. ウイルス感染細胞におけるウイルス増殖抑制効果の検討
ウイルス感染細胞を各エキスを含む培地で培養することで、各エキスのウイルス増殖抑制効果を検討した。2倍希釈のエキスを含む培地(2倍エキス含培地)での培養により、全てのエキスでH5N1(山口株)ウイルスのウイルス力価が検出限界(1.5 log10)以下まで低下した。5倍エキス含培地での培養では、ハスカップでのみウイルス力価が検出限界(1.5 log10)以下まで低下したが、コケモモ、マタタビでは,有意なウイルス力価の低下はみられなかった。
【0044】
【表3】
【0045】
ウイルス感染後の細胞にエキスを処理した実験では、2倍(ハスカップのみ5倍)希釈までのエキスを含む培地での培養によってウイルスの増殖が完全に抑えられていたことから、各エキスにはウイルスと混合することでウイルスを直接不活化するだけではなく、細胞に感染して細胞内に入り込んだウイルスに対しても増殖を抑制する効果があることが確認できた。
【0046】
5. 蛍光抗体法による細胞内のウイルス蛋白の検出
4.においてウイルス増殖が抑えられていた細胞内でのウイルス蛋白合成について、蛍光抗体法により検証した。結果を図2に示す。4.においてウイルス増殖抑制効果がみられたハスカップの5倍希釈のエキスを含むエキス含培地により培養した細胞では、ウイルス蛋白が全く検出されなかった。一方,他のエキスの5倍希釈ではウイルス対照と同程度のウイルス蛋白が検出された。
【0047】
6. Real-time RT-PCRによる細胞内のウイルス遺伝子の検出
4.において,ハスカップおよびコケモモエキス含培地(5倍希釈)により培養した細胞内でのvRNAおよびvmRNA合成について、Real-time RT-PCRを用いて解析した。結果を図3に示す。得られたCt は、MDCK細胞由来のハウスキーピング遺伝子であるGAPDHのCtとの相対比で表すことにより補正した(グラフの縦軸)。エキス不含培地で培養した細胞では、vRNAおよびvmRNAについてのCtが培養0時間から24時間まで継時的に低下し、vRNAおよびvmRNA合成が進行していることが確認された。一方、ハスカップエキス 含培地で培養した細胞では、vRNA、vmRNAともに培養1時間まではエキス不含培地で培養した細胞と同程度のCtであったが、培養3時間および6時間では、エキス不含培地で培養した細胞と比べ高いCtであった。しかし、培養24時間ではエキス不含培地で培養した細胞と同程度までCtが低下した。
【0048】
コケモモエキス含培地で培養した細胞では、vRNA、vmRNAともに培養1時間まではエキス不含培地で培養した細胞と比べCtが大きく低下した。培養3時間から24時間まではエキス不含培地で培養した細胞と比べやや高い値を示しつつ低下した(データ示さず)。
【0049】
感染細胞内でのウイルス動態を知る目的で、蛍光抗体法及びreal-time RT-PCRを用いて解析を行った。蛍光抗体法の結果から、ハスカップエキス存在下での培養では、感染細胞中でウイルス蛋白の合成が進行していないことが示された。また,real-time RT-PCRによって、ハスカップエキス含培地で培養・維持された感染細胞中でウイルスRNAの転写の進行が抑制されていることが示された。しかし,24時間後では,ウイルス対照のCt値と差が認められなかったことから,ウイルスRNAの産生は抑制されているものの,完全には阻害されていないと思われる。一方,ウイルス蛋白が合成されていないことから,ウイルス遺伝子からの蛋白質の翻訳が何らかのメカニズムで阻害されていることが示唆された。
【0050】
7. エキスによるMDCK細胞への細胞障害性の検討(LDH assay)
MDCK細胞への細胞障害性について、LDH assayにより検討した。結果を図4に示す。原液から8倍希釈まで、全てのエキスにおいて細胞障害性は認められなかった。
【0051】
果実エキスは、少なくとも原液から8倍濃度までの範囲の濃度では細胞障害性を全く示さなかった。さらに、5倍希釈のエキスにより2日間培養した細胞は、その後問題なく継代することができた(データ示さず)。これらのことから、果実エキスは、細胞に対して影響を与ることなく、細胞中のウイルスの複製に対してのみ影響を及ぼしていたことになる。
【産業上の利用可能性】
【0052】
本発明は、インフルエンザウイルスによる感染治療及び予防の分野に有用である。
【特許請求の範囲】
【請求項1】
ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤。
【請求項2】
前記水抽出物は、前記植物果実の粉砕または裁断物の水抽出物である請求項1に記載の抗インフルエンザウイルス剤。
【請求項3】
前記水抽出物は、熱水抽出物である請求項1または2に記載の抗インフルエンザウイルス剤。
【請求項4】
前記水抽出物に対する希釈剤をさらに含有する請求項1に記載の抗インフルエンザウイルス剤。
【請求項5】
インフルエンザ治療薬として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項6】
インフルエンザ感染予防薬として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項7】
インフルエンザ感染予防用組成物として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項8】
抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法であって、
前記植物果実の粉砕または裁断物を水中で煮沸し、水抽出液を固形分から分離することを含む、前記製造方法。
【請求項9】
水抽出液の固形分からの分離は、ろ過及び遠心分離の少なくとも一方を用いる、請求項8に記載の製造方法。
【請求項10】
前記水抽出液を乾燥して水抽出物の乾燥品を得る工程をさらに含む、請求項8または9に記載の製造方法。
【請求項1】
ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物を有効成分として含有する抗インフルエンザウイルス剤。
【請求項2】
前記水抽出物は、前記植物果実の粉砕または裁断物の水抽出物である請求項1に記載の抗インフルエンザウイルス剤。
【請求項3】
前記水抽出物は、熱水抽出物である請求項1または2に記載の抗インフルエンザウイルス剤。
【請求項4】
前記水抽出物に対する希釈剤をさらに含有する請求項1に記載の抗インフルエンザウイルス剤。
【請求項5】
インフルエンザ治療薬として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項6】
インフルエンザ感染予防薬として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項7】
インフルエンザ感染予防用組成物として用いるための請求項1に記載の抗インフルエンザウイルス剤。
【請求項8】
抗インフルエンザウイルス剤として用いるための、ハスカップ、コケモモ及びマタタビから成る群から選ばれる少なくとも1種の植物果実の水抽出物の製造方法であって、
前記植物果実の粉砕または裁断物を水中で煮沸し、水抽出液を固形分から分離することを含む、前記製造方法。
【請求項9】
水抽出液の固形分からの分離は、ろ過及び遠心分離の少なくとも一方を用いる、請求項8に記載の製造方法。
【請求項10】
前記水抽出液を乾燥して水抽出物の乾燥品を得る工程をさらに含む、請求項8または9に記載の製造方法。
【図1】


【図2】

【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2013−112644(P2013−112644A)
【公開日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願番号】特願2011−260579(P2011−260579)
【出願日】平成23年11月29日(2011.11.29)
【出願人】(504300088)国立大学法人帯広畜産大学 (96)
【Fターム(参考)】
【公開日】平成25年6月10日(2013.6.10)
【国際特許分類】
【出願日】平成23年11月29日(2011.11.29)
【出願人】(504300088)国立大学法人帯広畜産大学 (96)
【Fターム(参考)】
[ Back to top ]