
生体試料中のL−トリプトファン分析方法およびそれに用いるキット
【課題】L-トリプトファン特異的な酵素を用い、他のアミノ酸共存下でもL-トリプトファンを定量できる方法、この方法に利用できるキットおよび酵素センサーを提供する。
【解決手段】検体とL-トリプトファンオキシダーゼと水を混合する工程、得られた反応液を酸素の存在下に所定時間放置する工程、放置後の反応液中に存在する前記酵素の作用による反応生成物を計測する工程を含む、L-トリプトファンの定量方法。前記L-トリプトファンオキシダーゼは、所定アミノ酸配列を有し、かつ酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない。上記L-トリプトファンオキシダーゼを含むL-トリプトファンの定量用キット。上記L-トリプトファンオキシダーゼを用いる酵素センサー。
【解決手段】検体とL-トリプトファンオキシダーゼと水を混合する工程、得られた反応液を酸素の存在下に所定時間放置する工程、放置後の反応液中に存在する前記酵素の作用による反応生成物を計測する工程を含む、L-トリプトファンの定量方法。前記L-トリプトファンオキシダーゼは、所定アミノ酸配列を有し、かつ酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない。上記L-トリプトファンオキシダーゼを含むL-トリプトファンの定量用キット。上記L-トリプトファンオキシダーゼを用いる酵素センサー。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、生体試料中のL-トリプトファンを定量可能な生成物に変換し、その生成物の検出または定量によりL-トリプトファンの含有量を測定するのに適したL-トリプトファンオキシダーゼを用いるL-トリプトファン分析方法およびそれに用いるキットに関するものである。
【背景技術】
【0002】
血中をはじめとする生体試料中のアミノ酸濃度の定量は様々な疾病検出のマーカーとなり、その定量法の開発は医療的見地から強く望まれている。アミノ酸濃度の酵素的定量法は、その迅速さ・簡便さなど機器分析より優れた特長を有し、実際の医療現場で各種疾病検出を行うのに適した手法とされる。
【0003】
L-トリプトファンを酵素学的に定量する従来技術としては、(i)L-トリプトファンモノオキシゲナーゼ(非特許文献A、非特許文献B)、および(ii)L-アミノ酸オキシダーゼを用いる方法(特許文献A)が知られている。
【先行技術文献】
【特許文献】
【0004】
【特許文献A】特開2001-069974号公報
【非特許文献】
【0005】
【非特許文献A】Emanuele, J. J., Heasley, C. J., and Fitzpatrick, P. F. (1995) Arch Biochem Biophys 316, 241-248
【非特許文献B】Simonian, A. L., Rainina, E. I., Fitzpatrick, P. F., and Wild, J. R. (1997) Biosens Bioelectron 12, 363-371
【非特許文献C】Balibar, C. J., and Walsh, C. T. (2006) Biochemistry 45, 15444-15457
【非特許文献D】Onaka, H., Taniguchi, S., Igarashi, Y., and Furumai, T. (2002) J Antibiot 55, 1063-1071
【非特許文献E】Tonismagi, K., Samel, M., Trummal, K., Ronnholm, G., Siigur, J., Kalkkinen, N., and Siigur, E. (2006) Toxicon 48, 227-237
【非特許文献F】Tan, N. H., and Saifuddin, M. N. (1991) Int J Biochem 23, 323-327
【非特許文献G】Yang, H., Johnson, P. M., Ko, K. C., Kamio, M., Germann, M. W., Derby, C. D., and Tai, P. C. (2005) J Exp Biol 208, 3609-3622
【非特許文献H】Ehara, T., Kitajima, S., Kanzawa, N., Tamiya, T., and Tsuchiya, T. (2002) FEBS Lett 531, 509-512
【非特許文献I】Geueke, B., and Hummel, W. (2002) Enzyme Microb Technol 31, 77-87
【非特許文献J】Tong, H., Chen, W., Shi, W., Qi, F., and Dong, X. (2008) J Bacteriol 190, 4716-4721
【非特許文献K】Onaka, H., Taniguchi, S., Ikeda, H., Igarashi, Y., and Furumai, T. (2003) J Antibiot 56, 950-956
【発明の概要】
【発明が解決しようとする課題】
【0006】
上記の(i)L-トリプトファンモノオキシゲナーゼを用いる方法では、L-トリプトファン酸化に伴う酸素減少量を検出する必要がある。そのため、原理的に複雑な装置が求められる(非特許文献B)。また上記L-トリプトファンモノオキシゲナーゼは、L-トリプトファンへの活性を100%としたとき、L-フェニルアラニンに対し83%、L-メチオニンに対し42%という高い相対活性を示す(非特許文献A)。よってこの酵素を用いる方法は、簡便・安価かつL-トリプトファン特異性の高い検出を行うのには不適である。
【0007】
上記の(ii)L-アミノ酸オキシダーゼを用いる方法に関しては、公知のL-アミノ酸オキシダーゼは、一般的に基質特異性が低く、L-トリプトファン以外のL-アミノ酸複数種も反応基質とする。そのため、夾雑物存在下でL-トリプトファン特異的な検出をL-アミノ酸オキシダーゼにより行った例は知られていない。例えば、L-アミノ酸オキシダーゼとしては、ヘビ毒中の酵素が複数知られている。しかし、いずれの酵素も幅広いアミノ酸種に対し活性を有している(非特許文献E、F)。またこれ以外にも、軟体動物や細菌由来のL-アミノ酸オキシダーゼの報告例もある。しかし、これらも同様に基質特異性は広いことが知られており(非特許文献G, H, I, J)、これらの酵素を用いてL-トリプトファンを定量することは原理的に不可能である。
【0008】
比較的L-トリプトファンへの基質特異性が高い酵素としては、担子菌Coprinus sp.由来のL-アミノ酸オキシダーゼの報告例がある(特許文献A)。ただし上記酵素は、L-フェニルアラニンなど他の種類のアミノ酸にも7%程度の反応性を示す。従って、依然として夾雑物の多い生体試料の測定には不適である。また上記酵素は、L-トリプトファンへのKm値が650 μMであり、血中で数十μMオーダーの濃度でしか存在しないL-トリプトファンを正確に定量するには基質親和性が低いことも問題となる。実際にL-アミノ酸オキシゲナーゼを用いて試料中のL-トリプトファンを定量した例はこれまでに知られていない。さらに上記酵素は遺伝子同定されておらず、担子菌の長期(約1ヶ月)に渡る培養および複雑な精製過程がその製造には必要となる。以上の理由から、上記酵素による生体試料中のL-トリプトファン定量、および大量の酵素を要する実用化は困難であった。
【0009】
そこで本発明では、他のアミノ酸が共存する生体試料中においてもL-トリプトファン特異的な定量に応用可能な新たな酵素を探索し、この酵素を用いる、L-トリプトファンの新たな分析方法を提供することを目的とする。
【0010】
さらに本発明は、上記酵素的分析法を実施する際に利用できる測定用のキットを提供することも目的とする。加えて本発明は、上記酵素的分析法に利用できる酵素センサーを提供することも目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、細菌のビスインドール系抗生物質生合成経路中の遺伝子から、L-トリプトファンに基質特異性の高いL-トリプトファンオキシダーゼを発見した。この酵素を生体試料中に含まれるL-トリプトファンと反応させることにより検出可能な化合物が生成すること、さらに、この酵素が生体試料中に混在する他のアミノ酸に影響を受けずL-トリプトファン特異的な定量が可能であることを見出し、本発明を完成した。
【0012】
本発明は、以下に示す通りである。
[1]
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの分析方法であって、
前記L-トリプトファンオキシダーゼが、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記方法。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[2]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[1]に記載の方法。
[3]
工程(A)において検体との混合に用いられる前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤の存在下で保存されているものである、[1]または[2]に記載の方法。
[4]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[3]に記載の方法。
[5]
前記工程(C)で確認または計測する反応生成物が過酸化水素である[1]〜[4]のいずれかに記載の方法。
[6]
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファンオキシダーゼ
但し、前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[7]
前記(K1)L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[6]に記載のキット。
[8]
前記(K1)L-トリプトファンオキシダーゼは、前記酵素の安定化剤との混合物である、[6]または[7]に記載のキット。
[9]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[8]に記載のキット。
[10]
(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5)インドールピルビン酸検出薬の少なくとも一つをさらに含む、[6]〜[9]のいずれかに記載のキット。
[11]
L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物であって、
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記組成物。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[12]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[11]に記載の組成物。
[13]
前記酵素の安定化剤を含有する、[11]または[12]に記載の組成物。
[14]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[13]に記載の組成物。
[15]
L-トリプトファンオキシダーゼを用いるL-トリプトファンの検出または定量用酵素センサーであって、
前記検出用電極は過酸化水素検出用電極であり、かつ
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記センサー。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[16]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[15]に記載の酵素センサー。
[17]
前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に用いられる、[15]または[16]に記載の酵素センサー。
[18]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[17]に記載の酵素センサー。
[19]
過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極である[15]〜[18]のいずれかに記載の酵素センサー。
【発明の効果】
【0013】
本発明によれば、L-トリプトファンに対して特異的なトリプトファンオキシダーゼを用いることで、他のアミノ酸など多くの夾雑物を含む試料においても、L-トリプトファン特異的に迅速・簡便な検出を行うことができる。特に、血漿、血清または尿のような生体試料に対し本発明は有効であり、ペルオキシダーゼ等の酵素とカップリングさせることにより発色法や蛍光法でL-トリプトファンを定量できるのみならず、電極型酵素センサーを提供することもできる。
【図面の簡単な説明】
【0014】
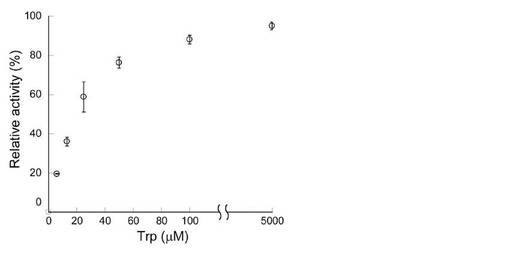
【図1】各L-トリプトファン濃度におけるStaOの活性を示す。
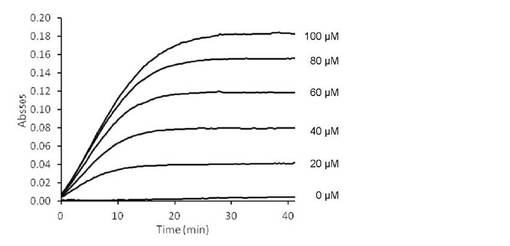
【図2】StaOによるL-トリプトファン検量線作成サンプルの吸光度の経時的変化を示す。
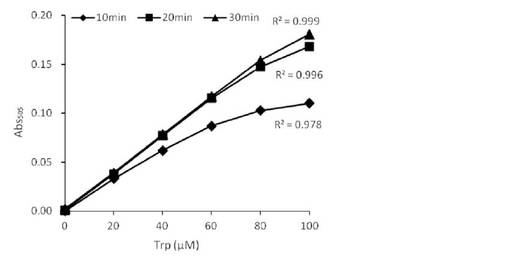
【図3】StaO使用サンプルにより作成したL-トリプトファン検量線を示す。
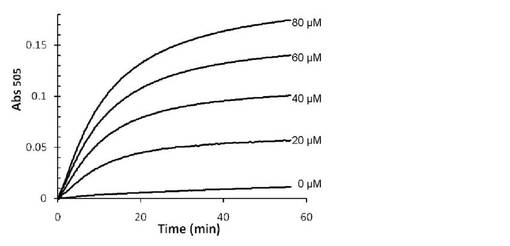
【図4】VioAによるL-トリプトファン検量線作成サンプルの吸光度の経時的変化を示す。
【図5】VioA使用サンプルにより作成したL-トリプトファン検量線を示す。
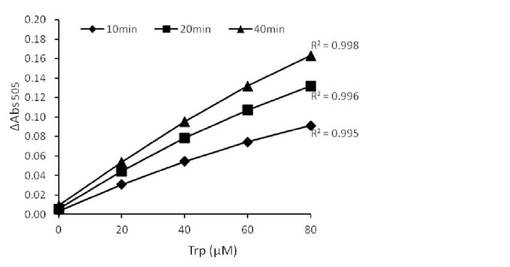
【図6】機器分析もしくはL-トリプトファンオキシダーゼによるヒト血漿試料中L-トリプトファン定量結果を示す。
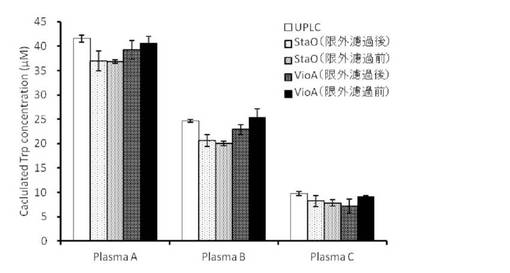
【図7】各種グリセロール濃度存在下におけるStaO残存活性の経時的変化を示す。
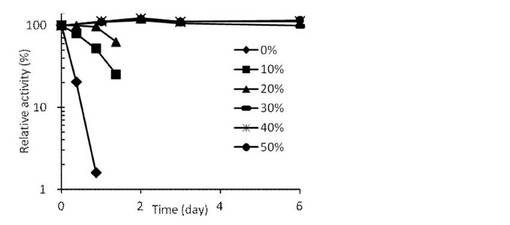
【図8】各種グリセロール濃度存在下におけるVioA残存活性の経時的変化を示す。
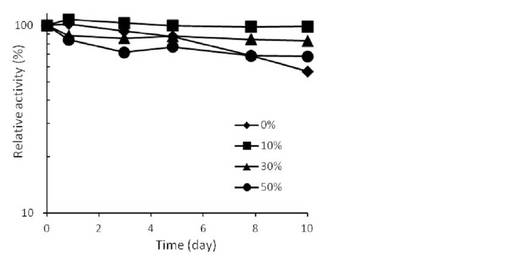
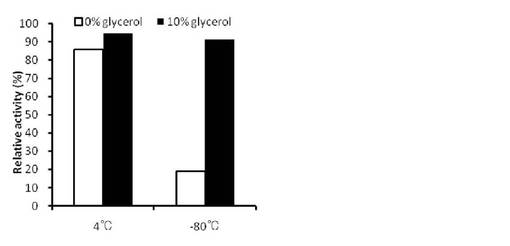
【図9】10%グリセロール存在下または非存在下における、4℃または-80℃で一晩VioAを保存した後の残存活性を示す。
【発明を実施するための形態】
【0015】
<L-トリプトファンの分析方法>
本発明のL-トリプトファンの分析方法は、
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含み、これらの工程を経ることで、検体中のL-トリプトファンの存在を確認するか、またはを定量する方法である。
【0016】
本発明の方法で、検体として用いられる生体試料は、L-トリプトファンを含む可能性のある試料であれば、如何なるものでもよい。生体試料にL-トリプトファンオキシダーゼを作用させて生じる、どの生成物を確認または定量することで生体試料中のL-トリプトファンを確認またはその濃度を測定するのかにより、生体試料は適宜選択することができる。例えば、発色剤や蛍光剤を利用して上記生成物を確認または定量する場合には無色の水溶液であることが好ましく、血清及び血漿などが例として挙げられる。
【0017】
本発明に用いる前記L-トリプトファンオキシダーゼは、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素である。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【0018】
(1)配列表の配列番号1に記載のアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素は、クロモバクテリウム・ビオラセウム(Chromobacterium violaceum)由来酵素であるVioAである。VioAは、同菌の生産する抗生物質violaceinの生合成経路の一部を触媒する酵素として同定され、L-トリプトファン酸化活性を有することが報告されていた酵素である(非特許文献C)。しかし、本酵素の基質特異性をはじめとする酵素学的性質については、これまでに検討されておらず、従って、VioAが、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることは、本発明において初めて明らかになったことである。さらに、VioAをL-トリプトファンの定量に用いることを報告した例はこれまでになかった。
【0019】
(1)配列表の配列番号2に記載のアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素は、ストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724由来酵素であるStaOである。StaOに関しては、同菌の生産する抗生物質staurosporineの生合成遺伝子クラスター中にその遺伝子が見出された酵素である(非特許文献D)。本遺伝子は上記VioAの遺伝子と有意な相同性を有しておらず、また遺伝子産物を生化学的に解析した例はないことから、その遺伝子産物の性状については未知のままであった。従って、StaOが、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることは、本発明において初めて明らかになったことである。
【0020】
尚、本願明細書において、「L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲である」における「L-フェニルアラニンに対するオキシダーゼ活性」及び「L-トリプトファンに対するオキシダーゼ活性」は、実施例における「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法においてタンパク質構成アミノ酸としてL-フェニルアラニン及びL-トリプトファンをそれぞれ用いた場合に得られる相対活性を意味する。上記「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法で得られる活性値は、何れも検出限界が0.5%である。また、本発明で用いるL-トリプトファンオキシダーゼは、分析精度が高まるという観点から、L-フェニルアラニンに対するオキシダーゼ活性がL-トリプトファンに対するオキシダーゼ活性の0〜2%の範囲であることが好ましく、0〜1.5%の範囲であることがより好ましく、0〜1%の範囲であることがさらに好ましい。
【0021】
さらに「L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない」における「L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対するオキシダーゼ活性」は、実施例における「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法においてタンパク質構成アミノ酸としてL-トリプトファン及びL-フェニルアラニン以外のアミノ酸をそれぞれ用いた場合に得られる相対活性を意味する。さらに、「オキシダーゼ活性を有さない」は、相対活性が、1%以下であること、好ましくは検出限界(0.5%)を超えた相対活性を示さないことを意味する。
【0022】
尚、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有すること、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さないことは、それぞれ実施例に記載の分析方法(定量方法)を用いることにより確認できる。また、タンパク質構成アミノ酸とは、L-チロシン、L-アラニン、L-システイン、L-アスパラギン酸、L-グルタミン酸、L-グリシン、L-ヒスチジン、L-イソロイシン、L-リジン、L-ロイシン、L-メチオニン、L-アスパラギン、L-プロリン、L-グルタミン、L-アルギニン、L-セリン、L-スレオニン、L-バリン、L-を意味する。
【0023】
本明細書で言う「1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から50個」の範囲は、欠失等を有するタンパク質が、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることを意味する。前記「1から50個」の範囲は、前記オキシダーゼ活性を有するタンパク質である割合が高いという観点から、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、さらに好ましくは1から10個、一層好ましくは1から7個、さらに一層好ましくは1から5個、特に好ましくは1から3個程度であることができる。
【0024】
本明細書で言う「配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列」における同一性は、前記アミノ酸配列の同一性を有するタンパク質が、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素である限り、特に限定されない。前記アミノ酸配列の同一性は、90%以上であれば特に限定されないが、好ましくは95%以上、さらに好ましくは96%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0025】
本発明で用いるL-トリプトファンオキシダーゼは、特定の種より生産されるものに限らず、特異的にL-トリプトファンのアミノ基を酸化的脱アミノ化するのに伴い過酸化水素を生産する酵素を意味する。
【0026】
さらに、本発明に用いるL-トリプトファンオキシダーゼは、同様の活性を有するものであれば、自然界より分離された生物に由来するもの、本酵素をコードする遺伝子を大腸菌や他の生物を宿主として発現させて得られる酵素またはタンパク質も含まれる。
【0027】
また、異種発現による生産法としては、例えば、同様の活性を有する生物種より抽出したゲノムDNAから該当する遺伝子をPCRにて増幅しpETもしくはpUCなどに組み込んだプラスミドベクターを構築したのち、BL21、JM109などの宿主菌株に形質転換し、培養する方法が挙げられる。これら以外の公知の方法も適宜用いることができる。
【0028】
本発明で用いるL-トリプトファンオキシダーゼの取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組換え技術により作製した組換えタンパク質でもよい。組換えタンパク質を作製する場合には、後述するように当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、上記L-トリプトファンオキシダーゼを産生することができる。
【0029】
上記L-トリプトファンオキシダーゼは、上記L-トリプトファンオキシダーゼをコードする遺伝子をベクター上に搭載し、このベクターによって宿主細胞を形質転換した後、形質転換させた宿主細胞を培養して培養物中に前記遺伝子がコードするタンパク質を蓄積し、蓄積したタンパク質を収集することを含む、生産方法により調製することができる。
【0030】
上記L-トリプトファンオキシダーゼをコードする遺伝子の取得方法は特に限定されない。本発明のL-トリプトファンオキシダーゼをコードする遺伝子は、例えば、配列番号1若しくは2に記載のアミノ酸配列または配列番号3若しくは4に記載した塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0031】
例えば、配列表の配列番号3または4に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、公知の方法を用いて行うことができる。
【0032】
本明細書中の配列表の配列番号1若しくは2に記載したアミノ酸配列または配列番号3若しくは4に示す塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いてクロモバクテリウム・ビオラセウム(Chromobacterium violaceum)NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノムライブラリーをスクリーニングすることにより本発明の遺伝子を単離することができる。ゲノムライブラリーは、クロモバクテリウム・ビオラセウム(Chromobacterium violaceum) NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724から常法により作製することができる。
【0033】
PCR法により本発明のL-トリプトファンオキシダーゼをコードする遺伝子を取得することもできる。上記クロモバクテリウム・ビオラセウム(Chromobacterium violaceum) NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノムライブラリーを鋳型として使用し、配列番号3若しくは4に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌(E. coli)等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0034】
上記したプローブ又はプライマーの調製、ゲノムライブラリーの構築、ゲノムライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知の方法に準じて適宜行うことができる。
【0035】
上記L-トリプトファンオキシダーゼの遺伝子は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製するベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、ベクターは発現ベクターである。発現ベクターにおいて上記遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモータは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0036】
細菌細胞で作動可能なプロモータとしては、バチルス・ステアロテルモフィルス・マルトジェニック・アミラーゼ遺伝子(Geobacillus stearothermophilus maltogenic amylase gene)、バチルス・リケニホルミスαアミラーゼ遺伝子(Bacillus licheniformis alpha-amylase gene)、バチルス・アミロリケファチエンス・BANアミラーゼ遺伝子(Bacillus amyloliquefaciens BAN amylase gene)、バチルス・サブチリス・アルカリプロテアーゼ遺伝子(Bacillus subtilis alkaline protease gene)もしくはバチルス・プミルス・キシロシダーゼ遺伝子(Bacillus pumilus xylosidase gene)のプロモータ、またはファージ・ラムダのPR若しくはPLプロモータ、大腸菌(E. coli)のlac、trp若しくはtacプロモータなどが挙げられる。
【0037】
哺乳動物細胞で作動可能なプロモータの例としては、SV40プロモータ、MT−1(メタロチオネイン遺伝子)プロモータ、またはアデノウイルス2主後期プロモータなどがある。昆虫細胞で作動可能なプロモータの例としては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュウロウイルス即時型初期遺伝子1プロモータ、またはバキュウロウイルス39K遅延型初期遺伝子プロモータ等がある。酵母宿主細胞で作動可能なプロモータの例としては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータなどが挙げられる。糸状菌細胞で作動可能なプロモータの例としては、ADH3プロモータまたはtpiAプロモータなどがある。
【0038】
また、上記L-トリプトファンオキシダーゼの遺伝子は必要に応じて、適切なターミネータに機能的に結合されてもよい。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターは更に、ポリアデニレーションシグナル(例えばSV40またはアデノウイルス5E1b領域由来のもの)、転写エンハンサ配列(例えばSV40エンハンサ)などの要素を有していてもよい。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターは更に、該ベクターが宿主細胞内で複製することを可能にするDNA配列を具備してもよく、その一例としてはSV40複製起点(宿主細胞が哺乳類細胞のとき)が挙げられる。
【0039】
L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、ジヒドロ葉酸レダクターゼ(DHFR)またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、または例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン若しくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。L-トリプトファンオキシダーゼの遺伝子、プロモータ、および所望によりターミネータおよび/または分泌シグナル配列をそれぞれ連結し、これらを適切なベクターに挿入する方法は当業者に周知である。
【0040】
L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターを導入される宿主細胞は、L-トリプトファンオキシダーゼの遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌および高等真核細胞等が挙げられる。
【0041】
細菌細胞の例としては、バチルスまたはストレプトマイセス等のグラム陽性菌又は大腸菌(E. coli)等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、または公知の方法でコンピテント細胞を用いることにより行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も公知であり、例えば、エレクトロポーレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0042】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0043】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0044】
昆虫細胞を宿主として用いる場合には、公知の方法を用いて組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる。
【0045】
バキュロウイルスとしては、例えば、ヨトウガ科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、Spodoptera frugiperdaの卵巣細胞であるSf9、Sf21〔バキュロウイルス・エクスプレッション・ベクターズ、ア・ラボラトリー・マニュアル、ダブリュー・エイチ・フリーマン・アンド・カンパニー(W. H. Freeman and Company)、ニューヨーク(New York)、(1992)〕、Trichoplusia niの卵巣細胞であるHiFive(インビトロジェン社製)等を用いることができる。
組換えウイルスを調製するための、昆虫細胞への組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法又はリポフェクション法等を挙げることができる。
【0046】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明で用いるL-トリプトファンオキシダーゼを単離精製するには、通常のタンパク質の単離、精製法を用いればよい。例えば、本発明で用いるL-トリプトファンオキシダーゼが、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のタンパク質の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、本発明のL-トリプトファンオキシダーゼを精製標品として得ることができる。
【0047】
L-トリプトファンオキシダーゼによるL-トリプトファンの酸化反応を以下の反応式Aに示す。
【化1】

【0048】
本発明で用いるL-トリプトファンオキシダーゼとは、上記式Aに示す反応を触媒する酵素を意味し、従って、その由来やアミノ酸配列の違いで限定されることはない。
【0049】
工程(A)
工程(A)におけるL-トリプトファンオキシダーゼの混合量は、10 mU/ml(トリプトファン1 μmolを1分間で消費する活性を1 Uとする)以上とすることが適当であり、水の混合量は、サンプル中のTrp濃度または反応系中の全質量に応じて適宜決定できるが、例えば、反応系中の全質量の5〜95%の範囲とすることができ、モル比としては、1モルのTrpに対して、例えば、1モル以上の水が存在すればよい。酵素反応を水溶液中で行う場合には過剰量の水が反応系中に存在することになる。L-トリプトファンオキシダーゼの混合量の上限は特にないが、実用的には、例えば、100 mU/ml以下であることができる。しかし、L-トリプトファンオキシダーゼの混合量および水の混合量は、この範囲に限定する意図ではなく、適宜調整できる。
【0050】
さらに、L-トリプトファンオキシダーゼおよび水に加えて、好ましくは、L-トリプトファンオキシダーゼの至適pHを考慮したpHを示す緩衝液を含むことができる。緩衝液を用いる場合には、水は緩衝液として供給することもできる。さらに、L-トリプトファンオキシダーゼの安定化剤をさらに含むこともできる。L-トリプトファンオキシダーゼの安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0051】
工程(B)
工程(B)では、前記混合により得られた反応液を酸素の存在下に所定時間放置する。
L-トリプトファンオキシダーゼによるL-トリプトファン酸化反応においては、反応式Aに示すように、L-トリプトファン脱アミノ化生成物であるインドール-3-ピルビン酸と共に、アンモニア(NH3)と過酸化水素(H2O2)が生成物として得られる。上記反応を、例えば、空気と接触する状態で実施することで、反応液中の溶存酸素として上記酸素は供給される。反応液中酸素を供給する目的で反応液に空気などの酸素含有気体を強制的に供給する必要は通常はない。酵素反応に必要とされる酸素量が微量であり、溶存酸素により十分に賄えるためである。酵素のための放置時間は、例えば、使用する酵素量にもよるが、例えば、10分〜1時間の範囲とすることができる。しかし、この範囲に限定する意図ではなく、適宜調整できる。
【0052】
工程(C)
工程(C)では、放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または反応生成物の少なくとも一種の量を計測する。
存在確認または定量に用いられる生成物が過酸化水素である場合、例えばペルオキシダーゼ反応を用いて測定する方法等の公知の方法により、過酸化水素の存在確認または定量可能である。ペルオキシダーゼ反応を用いて測定する場合、使用可能なペルオキシダーゼは過酸化水素の存在確認または定量に利用可能な酵素であればよく、例えば西洋わさび由来ペルオキシダーゼが挙げられる。また、使用するペルオキシダーゼの基質となり得るものであれば発色剤として使用可能であり、西洋わさび由来ペルオキシダーゼを用いる場合には4-アミノアンチピリン:フェノールなどが挙げられる。西洋わさび由来ペルオキシダーゼを用いる過酸化水素の存在確認または定量のための反応は以下に示す通りである。
【0053】
【化2】
【0054】
4-アミノアンチピリン等の発色剤や蛍光剤は、使用されるペルオキシダーゼの種類によって適宜選択することが可能である。
【0055】
L-トリプトファンオキシダーゼ反応の生成物である過酸化水素は、過酸化水素電極を用いた電流検出型センサーを用いて測定することもできる。この測定により、過酸化水素の存在確認および定量の何れも可能である。過酸化水素電極としては、例えば、ペルオキシダーゼを牛血清アルブミンとともにグルタルアルデヒドに固定化した膜とフェロセンをカーボンペーストに含有させたものを電極として用いるセンサーを挙げることができる。
【0056】
存在確認または定量に用いられる生成物がアンモニアである場合には、アンモニア検出薬を用いて測定することができる。アンモニア検出薬としては、例えば、フェノールと次亜塩素酸の組み合わせによるインドフェノール法を挙げることができる。具体的には、サンプルをフェノール・ニトロプルシド溶液および過塩素酸溶液と混合して発色させることで存在確認ができ、さらに、この発色を635 nmの吸光度を測定することにより、アンモニア定量が可能である。
【0057】
存在確認または定量に用いられる生成物がL-トリプトファンの脱アミノ化生成物であるインドールピルビン酸である場合には、2-オキソ酸還元酵素を用いて、インドールピルビン酸の存在確認または定量を行うことができる。
【0058】
<L-トリプトファンの分析用キット>
本発明は、以下の試薬を含むL-トリプトファンの分析用キットを包含する。
(K1)L-トリプトファンオキシダーゼ
上記L-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。L-トリプトファンの分析には、L-トリプトファンの存在確認および定量を含む。
【0059】
上記L-トリプトファンオキシダーゼは、このL-トリプトファンオキシダーゼの安定化剤との混合物であることもできる。安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。安定化剤は、グリセロールであることが、酵素に対する安定化効果に優れるため好ましい。
【0060】
本発明のキットは、(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5) インドールピルビン酸検出薬の少なくとも一つをさらに含むことができる。
【0061】
(K2)反応用緩衝液は、反応液中を定量等の反応に適したpHに維持するために用いられる。後述の実施例に示すL-トリプトファンオキシダーゼは、至適pHが8〜9の範囲があるので、この範囲のpHを有する緩衝液であることが望ましい。
【0062】
(K3)過酸化水素検出用試薬は、過酸化水素の検出を、例えば、発色もしくは蛍光によって行う場合に用いる。過酸化水素検出用試薬としては、例えば、ペルオキシダーゼとその基質となり得る発色剤の組合せであることができる。具体的には、西洋わさびペルオキシダーゼと2-アミノアンチピリン・フェノールの組み合わせを挙げることができる。
【0063】
(K4)アンモニア検出薬としては、例えば、フェノールと次亜塩素酸の組み合わせによるインドフェノール法を挙げることができる。
【0064】
(K5)インドールピルビン酸検出薬としては、例えば、2-オキソ酸還元酵素を用いることができる。
【0065】
<L-トリプトファンの分析用組成物>
本発明は、L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物も包含する。この組成物に含まれるL-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。L-トリプトファンの分析には、L-トリプトファンの存在確認および定量を含む。本発明の分析用組成物は、前記L-トリプトファンに加えて、例えば、前記酵素の安定化剤、緩衝液または緩衝剤などを含有することができる。
【0066】
このL-トリプトファンオキシダーゼの安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。安定化剤は、グリセロールであることが、酵素に対する安定化効果に優れるため好ましい。
【0067】
酵素と安定化剤との混合比は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲であり、より好ましくは30〜50質量部の範囲である。
【0068】
<酵素センサー>
本発明は、L-トリプトファンオキシダーゼを用いたL-トリプトファンの検出または定量用酵素センサーを包含する。この酵素センサーに用いるL-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。
【0069】
前記検出用電極は過酸化水素検出用電極である。過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極であることができる。L-トリプトファンオキシダーゼがL-トリプトファンと反応することで、過酸化水素が生成するので、この過酸化水素を過酸化水素検出用電極で検出することができる。酵素式過酸化水素電極としては、例えば、ペルオキシダーゼを牛血清アルブミンとともにグルタルアルデヒドに固定化した膜とフェロセンをカーボンペーストに含有させたものを電極として用いるセンサーを挙げることができる。隔膜式過酸化水素電極は、隔膜により過酸化水素と反応する電極が隔離されたタイプの電極である。
【0070】
前記L-トリプトファンオキシダーゼは、L-トリプトファンの検出または定量をより高精度に行うという観点から、検出用電極の表面または検出用電極の近傍に配置されることが好ましく、検出用電極の表面に配置される場合には、検出用電極の表面に固定化されても固定化されなくてもよい。検出用電極の表面に固定化されることで、本発明のセンサーを繰り返し利用できる利点はある。さらに、前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置されることもできる。前記安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【実施例】
【0071】
以下に実施例を挙げて本発明を具体的に説明するが本発明はこれに限定されるものではない。
【0072】
1.L-トリプトファンオキシダーゼの調製例
クロモバクテリウム・ビオラセウム(Chromobacterium violaceum)NBRC 12614ゲノムDNAを調製し、これを鋳型として塩基配列データベース上の各菌の配列(AF172851.1)を元に設計した配列番号5および6のプライマーを用いてPCRを行い、L-トリプトファンオキシダーゼ遺伝子vioAを増幅した。増幅産物をpET-28aに挿入し、vioA発現用プラスミドとした。また、ストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノム断片を含むコスミドpTYMCsta(非特許文献D)をStuI処理した断片を、HincIIで処理したベクターpTYM19(非特許文献K)に挿入し、L-トリプトファンオキシダーゼ遺伝子staOをサブクローニングした。Stratagene社製QuikChange site-directed mutagenesis kitと配列番号7〜10のプライマーを用い、上記サブクローニングプラスミドの配列を改変した。このプラスミドからNdeIおよびHindIII処理によりstaO遺伝子を切り出し、同制限酵素で処理したpET-26bに挿入し、staO発現用プラスミドとした。なお、各酵素のN末またはC末にHis-tagが付加されるようにプライマー設計を行った。各発現用プラスミドで大腸菌BL21 (DE3)株を形質転換し、VioAもしくはStaO異種発現株とした。プライマーの塩基配列は以下の通りである。
VioA-F: ATTCTAGACATATGAAGCATTCTTCCGATATCTG(配列番号5)
VioA-R: AATAAGCTTCGCGGCGATGCGCTG(配列番号6)
StaO-N-sense: TACTGGAGGAAACATATGACGGCACCC(配列番号7)
StaO-N-anti: CAAGGGTGCCGTCATATGTTTCCTCCA(配列番号8)
StaO-C-sense: GACCGGTCGGCGAAGCTTTCTTCGACCTG(配列番号9)
StaO-C-anti: GCAGGTCGAAGAAAGCTTCGCCGACCGGT(配列番号10)
【0073】
各異種発現株を37℃の振盪培養でOD600が0.6〜0.8に達するまで培養し、IPTGを終濃度0.5 mMになるように添加することで発現誘導をかけた。発現誘導後の培養は16℃で16時間行い、可溶性画分に目的酵素が得られた。またStaOに関しては、Takara社製シャペロンプラスミドpG-KJE8を共発現させると収量がさらに向上した。
【0074】
各発現株破砕液上清をGEヘルスケア社製Ni Sepharoseカラムにロードし、20 mM Tris-HCl, 50 mM imidazole (pH 8.0)溶液で洗浄後、20 mM Tris-HCl, 500 mM imidazole (pH 8.0)溶液で溶出させることによって目的酵素を精製・回収した。これらの酵素の塩基配列を決定し、配列番号3および4に示す。さらに酵素のアミノ酸配列を配列番号1および2に示す。
【0075】
2.L-トリプトファンオキシダーゼの活性測定条件例
20 mM トリス−塩酸バッファー (pH 8.0)、5 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、および任意の量のL-トリプトファンオキシダーゼ酵素液を含む反応液を調製し、30℃での505 nmの吸光度変化を測定する。キノンイミン色素のモル吸光係数は6.4 mM-1・cm-1とする。1分間に1 μmolの過酸化水素を生成する活性を1 Uと定義する。本法の検出限界は0.5%である。
【0076】
3.活性のpH依存性
リン酸カリウムバッファー (pH 6.5, 7.0, 7.5, 8.0, 8.5)およびトリス−塩酸バッファー (pH 8.0, 8.5, 9.0)をバッファーとして用い、各バッファー使用時のStaOの活性測定を行った。20 mMの上記のうちいずれかのバッファー、5 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、およびStaO酵素液を含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。その結果、StaOの酵素活性はpH8.0-9.0で最大になることが示された。尚、VioAの至適pHは非特許文献Cに9.25であることが報告されている。
【0077】
4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性
20 mM トリス−塩酸バッファー (pH 8.0)、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、0.2〜20 mU/ml L-トリプトファンオキシダーゼと0.5 mMのアミノ酸(20種類のタンパク構成アミノ酸のうちいずれか1種)を含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。得られた測定値から各アミノ酸との相対活性を算出し、表1のような結果となった。本法の検出限界は0.5%である。
【0078】
StaOはL-トリプトファン以外のアミノ酸とは反応性を示さず(相対活性は検出限界以下であった)、L-フェニルアラニンに対してのみ、L-トリプトファンの0.5%〜1%というごく低い相対活性が検出された。またVioAに関してはL-トリプトファン以外のアミノ酸への反応性は検出されなかった(相対活性は検出限界以下であった)。以上のように、StaOとVioAいずれに関しても、L-トリプトファンに対してきわめて高い基質特異性を有していることが示された。
【0079】
5.L-トリプトファンオキシダーゼのL-トリプトファンに対するKm値
20 mM トリス−塩酸バッファー (pH 8.0)、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、0.5 mU/ml L-トリプトファンオキシダーゼと、0, 6.3, 13, 25, 50, 100, 500μMのL-トリプトファンを含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。
【0080】
各L-トリプトファン濃度で算出されたStaOの活性を図1に示す。この結果から、StaOのL-トリプトファンへのKm値は19μMと算出された。またVioAのKm値に関しては、30μMであると報告されている(非特許文献C)。これらの値は、Coprinus sp.由来のL-アミノ酸オキシダーゼの値(650μM)と比べてきわめて低く、また一般的にアミノ酸定量用に用いられている酵素の基質に対するKm値と比べても同等以下に低い値である。このことは、StaOおよびVioAはL-トリプトファン濃度の低いサンプルにおいても定量用酵素として優れた性質を有することを示している。
【0081】
【表1】
【0082】
6.L-トリプトファンの検量線の作成
40 mM トリス−塩酸バッファー (pH 8.0)、2 mMフェノール、2 mM 4-アミノアンチピリン、30 U/ml西洋わさび由来ペルオキシダーゼ、20 mU/ml L-トリプトファンオキシダーゼ酵素液を含む反応液を調製した。これと等量の0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液を標準試料として混合した。30℃で反応を進行させ、30℃での505 nmの吸光度の経時的変化を測定し、反応の進行をモニタリングした。
【0083】
上述のように作製したサンプルの測定結果を図2〜5に示す。StaO, VioAいずれを用いた場合でも、遅くとも反応開始20分後以降はL-トリプトファン濃度と吸光度の間に良好な直線関係が見られ、本反応系で検量線が作成可能であることが示された。また特にStaOサンプルに関しては、反応時間を過剰に長くした場合でも吸光度上昇はほぼ止まるという変化を示した。こうした性質は、たとえ反応時間がばらついたとしても誤差の出にくい、定量用酵素として適した性質と言える。
【0084】
7.ヒト血漿サンプル中のL-トリプトファン定量
7−1.ヒト血漿試料の前処理
ヒト血漿試料はコージンバイオより購入し、−20℃で保管していた3検体を用いた。それぞれ使用直前に解凍し、そのまま、もしくはMicrocon YM-10で限外濾過による除タンパク処理後に、下記のL-トリプトファン定量に供した。
【0085】
7−2.機器分析によるヒト血漿試料中L-トリプトファンの定量
酵素法による定量の比較対象として、超高速アミノ酸分析システムを用いたプレカラム誘導体化法による定量を行った。前述のようにヒト血漿試料を除タンパク質処理したものを超高速アミノ酸分析システムWaters UPLC Amino Acid Analysis Solution systemに供した。サンプルの誘導体化、分析操作などは上記システムの説明書の手順に従った。0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液を標準試料として検量線を作製し、血漿試料中L-トリプトファンの定量を行った。
【0086】
7−3.L-トリプトファンオキシダーゼによるヒト血漿試料中L-トリプトファンの定量
40 mM トリス−塩酸バッファー (pH 8.0)、2 mMフェノール、2 mM 4-アミノアンチピリン、30 U/ml西洋わさび由来ペルオキシダーゼ、20 mU/ml L-トリプトファンオキシダーゼ酵素液を含む反応液を調製した。これと等量の0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液、もしくは除タンパク処理前後のヒト血漿試料を混合した。30℃で反応を進行させ、30分後に505 nmの吸光度を測定した。L-トリプトファン水溶液サンプルの測定値から検量線を作製し、血漿試料中L-トリプトファンの定量を行った。
【0087】
7−4.機器分析もしくはL-トリプトファンオキシダーゼによるヒト血漿試料の定量結果の比較
上記の測定結果を図6に示す。StaO, VioAいずれの酵素を使った定量系でも、機器分析による測定値に近い値が得られた。よってL-トリプトファンオキシダーゼによる定量法がヒト血漿試料においても有効であることが裏付けられた。
【0088】
一般的な酵素法による生体試料の分析では、あらかじめ除タンパク質処理を行わないと定量性が損なわれるという例が多い。この場合、生体試料の前処理として除タンパク処理が必要となる。一方StaO, VioAはいずれも、除タンパク質処理未処理の試料でも定量性に影響は見られなかった。よって、上記酵素による定量法には試料の前処理が不要であり、酵素法としても特に簡便かつ正確な定量法であることがわかった。
【0089】
8.安定性
StaO, VioA精製酵素液に終濃度0〜50% (v/v)となるようにグリセロールを添加した。各酵素液を4℃で保存し、経時的にサンプリングして活性測定を行った。各種グリセロール濃度存在下におけるStaOまたはVioAの残存活性の経時的変化を図7と図8に示す。StaOに関しては、バッファーのみを含む溶液中では不安定ではあるが、30%以上のグリセロール添加条件で高い安定性を示した。VioAはグリセロール非添加でも比較的安定であるが、10%以上のグリセロールを添加するとその安定性はさらに高まった。
【0090】
VioA精製酵素液に終濃度0または10% (v/v)となるようにグリセロールを添加した。各酵素液を4℃または-80℃で一晩保存した後、活性測定を行った。10%グリセロール存在下または非存在下におけるVioA酵素液の残存活性を図9に示す。VioAの凍結融解時には失活が見られたが、10% グリセロールの添加によりそのような失活は見られなくなった。
【0091】
以上のように、StaOはグリセロール 30%添加、VioAは非添加(凍結融解を行うときには10%添加)で安定な酵素液として使用可能であることが示された。
【0092】
9.反応温度依存性
反応温度20, 30, 40, 50, 60℃でのStaO, VioAの活性測定を行った。20 mM トリス−塩酸バッファー (pH 8.0)、1 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、およびL-トリプトファンオキシダーゼ酵素液を組成とする活性測定反応液を用いた。上記反応液のうちペルオキシダーゼとL-トリプトファンオキシダーゼ酵素液以外を混合し、上記各種温度で10分間プレインキュベーションを行った。上記反応液の温度を保ったままペルオキシダーゼとL-トリプトファンオキシダーゼ酵素液を添加して反応を開始させ、速やかに505 nmの吸光度変化を測定した。測定結果から算出した、各温度におけるStaO, VioAの相対活性値を表2に示す。両酵素とも、反応温度の上昇と共に活性の上昇が見られた。なお、StaOでは60℃、VioAでは50℃以上で、高温による失活の影響が顕著となり正確な活性測定が困難になった。
【0093】
【表2】
【0094】
10.熱安定性
20%グリセロールを含んだStaO酵素液、および10%グリセロールを含んだVioA酵素液を30, 40, 50, 60, 70℃で1時間熱処理した後、活性測定を行った。熱処理後の各酵素液の残存活性を表3に示す。いずれの酵素も40℃以上の熱処理では顕著な熱失活が見られ、50℃以上の熱処理では残存活性が検出限界以下となった。
【0095】
【表3】
【産業上の利用可能性】
【0096】
L-トリプトファンは、必須アミノ酸であり、極端に不足した場合、ナイアシン欠乏症状を示す事が風土病ペラグラとして知られている。また、過剰に摂取した場合、好酸球増加や筋肉痛症候群の原因になるか可能性が指摘されている。このため、L-トリプトファンの定量は、食品分析、医薬品・サプリメントの品質管理、過剰症・欠乏症時の血液検査及び酵素センサーとして利用が考えられる。また、代謝経路上、ナイアシンやNADなど生体内で重要な機能を持つ物質の原材料となることから、アミノグラムによる疾病診断「アミノインデックス」においても様々な疾病のバイオマーカーを構成する数値の一つとして活用できる。これらの理由から、食品や生体試料などについて、L-トリプトファン量を測定することは、産業的にも医学的にも重要な技術となると考えられるが、現状では、高速液体クロマトフラフィーをはじめとする非常に高価な機器及び試薬が必要となる方法しか実用化されていない。したがって、本発明は、L-トリプトファン定量用キットや酵素センサーなどの形態で商品化が可能であり、安価かつ簡便なL-トリプトファン定量法として事業化できる。
【技術分野】
【0001】
本発明は、生体試料中のL-トリプトファンを定量可能な生成物に変換し、その生成物の検出または定量によりL-トリプトファンの含有量を測定するのに適したL-トリプトファンオキシダーゼを用いるL-トリプトファン分析方法およびそれに用いるキットに関するものである。
【背景技術】
【0002】
血中をはじめとする生体試料中のアミノ酸濃度の定量は様々な疾病検出のマーカーとなり、その定量法の開発は医療的見地から強く望まれている。アミノ酸濃度の酵素的定量法は、その迅速さ・簡便さなど機器分析より優れた特長を有し、実際の医療現場で各種疾病検出を行うのに適した手法とされる。
【0003】
L-トリプトファンを酵素学的に定量する従来技術としては、(i)L-トリプトファンモノオキシゲナーゼ(非特許文献A、非特許文献B)、および(ii)L-アミノ酸オキシダーゼを用いる方法(特許文献A)が知られている。
【先行技術文献】
【特許文献】
【0004】
【特許文献A】特開2001-069974号公報
【非特許文献】
【0005】
【非特許文献A】Emanuele, J. J., Heasley, C. J., and Fitzpatrick, P. F. (1995) Arch Biochem Biophys 316, 241-248
【非特許文献B】Simonian, A. L., Rainina, E. I., Fitzpatrick, P. F., and Wild, J. R. (1997) Biosens Bioelectron 12, 363-371
【非特許文献C】Balibar, C. J., and Walsh, C. T. (2006) Biochemistry 45, 15444-15457
【非特許文献D】Onaka, H., Taniguchi, S., Igarashi, Y., and Furumai, T. (2002) J Antibiot 55, 1063-1071
【非特許文献E】Tonismagi, K., Samel, M., Trummal, K., Ronnholm, G., Siigur, J., Kalkkinen, N., and Siigur, E. (2006) Toxicon 48, 227-237
【非特許文献F】Tan, N. H., and Saifuddin, M. N. (1991) Int J Biochem 23, 323-327
【非特許文献G】Yang, H., Johnson, P. M., Ko, K. C., Kamio, M., Germann, M. W., Derby, C. D., and Tai, P. C. (2005) J Exp Biol 208, 3609-3622
【非特許文献H】Ehara, T., Kitajima, S., Kanzawa, N., Tamiya, T., and Tsuchiya, T. (2002) FEBS Lett 531, 509-512
【非特許文献I】Geueke, B., and Hummel, W. (2002) Enzyme Microb Technol 31, 77-87
【非特許文献J】Tong, H., Chen, W., Shi, W., Qi, F., and Dong, X. (2008) J Bacteriol 190, 4716-4721
【非特許文献K】Onaka, H., Taniguchi, S., Ikeda, H., Igarashi, Y., and Furumai, T. (2003) J Antibiot 56, 950-956
【発明の概要】
【発明が解決しようとする課題】
【0006】
上記の(i)L-トリプトファンモノオキシゲナーゼを用いる方法では、L-トリプトファン酸化に伴う酸素減少量を検出する必要がある。そのため、原理的に複雑な装置が求められる(非特許文献B)。また上記L-トリプトファンモノオキシゲナーゼは、L-トリプトファンへの活性を100%としたとき、L-フェニルアラニンに対し83%、L-メチオニンに対し42%という高い相対活性を示す(非特許文献A)。よってこの酵素を用いる方法は、簡便・安価かつL-トリプトファン特異性の高い検出を行うのには不適である。
【0007】
上記の(ii)L-アミノ酸オキシダーゼを用いる方法に関しては、公知のL-アミノ酸オキシダーゼは、一般的に基質特異性が低く、L-トリプトファン以外のL-アミノ酸複数種も反応基質とする。そのため、夾雑物存在下でL-トリプトファン特異的な検出をL-アミノ酸オキシダーゼにより行った例は知られていない。例えば、L-アミノ酸オキシダーゼとしては、ヘビ毒中の酵素が複数知られている。しかし、いずれの酵素も幅広いアミノ酸種に対し活性を有している(非特許文献E、F)。またこれ以外にも、軟体動物や細菌由来のL-アミノ酸オキシダーゼの報告例もある。しかし、これらも同様に基質特異性は広いことが知られており(非特許文献G, H, I, J)、これらの酵素を用いてL-トリプトファンを定量することは原理的に不可能である。
【0008】
比較的L-トリプトファンへの基質特異性が高い酵素としては、担子菌Coprinus sp.由来のL-アミノ酸オキシダーゼの報告例がある(特許文献A)。ただし上記酵素は、L-フェニルアラニンなど他の種類のアミノ酸にも7%程度の反応性を示す。従って、依然として夾雑物の多い生体試料の測定には不適である。また上記酵素は、L-トリプトファンへのKm値が650 μMであり、血中で数十μMオーダーの濃度でしか存在しないL-トリプトファンを正確に定量するには基質親和性が低いことも問題となる。実際にL-アミノ酸オキシゲナーゼを用いて試料中のL-トリプトファンを定量した例はこれまでに知られていない。さらに上記酵素は遺伝子同定されておらず、担子菌の長期(約1ヶ月)に渡る培養および複雑な精製過程がその製造には必要となる。以上の理由から、上記酵素による生体試料中のL-トリプトファン定量、および大量の酵素を要する実用化は困難であった。
【0009】
そこで本発明では、他のアミノ酸が共存する生体試料中においてもL-トリプトファン特異的な定量に応用可能な新たな酵素を探索し、この酵素を用いる、L-トリプトファンの新たな分析方法を提供することを目的とする。
【0010】
さらに本発明は、上記酵素的分析法を実施する際に利用できる測定用のキットを提供することも目的とする。加えて本発明は、上記酵素的分析法に利用できる酵素センサーを提供することも目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、細菌のビスインドール系抗生物質生合成経路中の遺伝子から、L-トリプトファンに基質特異性の高いL-トリプトファンオキシダーゼを発見した。この酵素を生体試料中に含まれるL-トリプトファンと反応させることにより検出可能な化合物が生成すること、さらに、この酵素が生体試料中に混在する他のアミノ酸に影響を受けずL-トリプトファン特異的な定量が可能であることを見出し、本発明を完成した。
【0012】
本発明は、以下に示す通りである。
[1]
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの分析方法であって、
前記L-トリプトファンオキシダーゼが、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記方法。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[2]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[1]に記載の方法。
[3]
工程(A)において検体との混合に用いられる前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤の存在下で保存されているものである、[1]または[2]に記載の方法。
[4]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[3]に記載の方法。
[5]
前記工程(C)で確認または計測する反応生成物が過酸化水素である[1]〜[4]のいずれかに記載の方法。
[6]
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファンオキシダーゼ
但し、前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[7]
前記(K1)L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[6]に記載のキット。
[8]
前記(K1)L-トリプトファンオキシダーゼは、前記酵素の安定化剤との混合物である、[6]または[7]に記載のキット。
[9]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[8]に記載のキット。
[10]
(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5)インドールピルビン酸検出薬の少なくとも一つをさらに含む、[6]〜[9]のいずれかに記載のキット。
[11]
L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物であって、
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記組成物。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[12]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[11]に記載の組成物。
[13]
前記酵素の安定化剤を含有する、[11]または[12]に記載の組成物。
[14]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[13]に記載の組成物。
[15]
L-トリプトファンオキシダーゼを用いるL-トリプトファンの検出または定量用酵素センサーであって、
前記検出用電極は過酸化水素検出用電極であり、かつ
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記センサー。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
[16]
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、[15]に記載の酵素センサー。
[17]
前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に用いられる、[15]または[16]に記載の酵素センサー。
[18]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[17]に記載の酵素センサー。
[19]
過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極である[15]〜[18]のいずれかに記載の酵素センサー。
【発明の効果】
【0013】
本発明によれば、L-トリプトファンに対して特異的なトリプトファンオキシダーゼを用いることで、他のアミノ酸など多くの夾雑物を含む試料においても、L-トリプトファン特異的に迅速・簡便な検出を行うことができる。特に、血漿、血清または尿のような生体試料に対し本発明は有効であり、ペルオキシダーゼ等の酵素とカップリングさせることにより発色法や蛍光法でL-トリプトファンを定量できるのみならず、電極型酵素センサーを提供することもできる。
【図面の簡単な説明】
【0014】
【図1】各L-トリプトファン濃度におけるStaOの活性を示す。
【図2】StaOによるL-トリプトファン検量線作成サンプルの吸光度の経時的変化を示す。
【図3】StaO使用サンプルにより作成したL-トリプトファン検量線を示す。
【図4】VioAによるL-トリプトファン検量線作成サンプルの吸光度の経時的変化を示す。
【図5】VioA使用サンプルにより作成したL-トリプトファン検量線を示す。
【図6】機器分析もしくはL-トリプトファンオキシダーゼによるヒト血漿試料中L-トリプトファン定量結果を示す。
【図7】各種グリセロール濃度存在下におけるStaO残存活性の経時的変化を示す。
【図8】各種グリセロール濃度存在下におけるVioA残存活性の経時的変化を示す。
【図9】10%グリセロール存在下または非存在下における、4℃または-80℃で一晩VioAを保存した後の残存活性を示す。
【発明を実施するための形態】
【0015】
<L-トリプトファンの分析方法>
本発明のL-トリプトファンの分析方法は、
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含み、これらの工程を経ることで、検体中のL-トリプトファンの存在を確認するか、またはを定量する方法である。
【0016】
本発明の方法で、検体として用いられる生体試料は、L-トリプトファンを含む可能性のある試料であれば、如何なるものでもよい。生体試料にL-トリプトファンオキシダーゼを作用させて生じる、どの生成物を確認または定量することで生体試料中のL-トリプトファンを確認またはその濃度を測定するのかにより、生体試料は適宜選択することができる。例えば、発色剤や蛍光剤を利用して上記生成物を確認または定量する場合には無色の水溶液であることが好ましく、血清及び血漿などが例として挙げられる。
【0017】
本発明に用いる前記L-トリプトファンオキシダーゼは、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素である。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【0018】
(1)配列表の配列番号1に記載のアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素は、クロモバクテリウム・ビオラセウム(Chromobacterium violaceum)由来酵素であるVioAである。VioAは、同菌の生産する抗生物質violaceinの生合成経路の一部を触媒する酵素として同定され、L-トリプトファン酸化活性を有することが報告されていた酵素である(非特許文献C)。しかし、本酵素の基質特異性をはじめとする酵素学的性質については、これまでに検討されておらず、従って、VioAが、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることは、本発明において初めて明らかになったことである。さらに、VioAをL-トリプトファンの定量に用いることを報告した例はこれまでになかった。
【0019】
(1)配列表の配列番号2に記載のアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない酵素は、ストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724由来酵素であるStaOである。StaOに関しては、同菌の生産する抗生物質staurosporineの生合成遺伝子クラスター中にその遺伝子が見出された酵素である(非特許文献D)。本遺伝子は上記VioAの遺伝子と有意な相同性を有しておらず、また遺伝子産物を生化学的に解析した例はないことから、その遺伝子産物の性状については未知のままであった。従って、StaOが、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることは、本発明において初めて明らかになったことである。
【0020】
尚、本願明細書において、「L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲である」における「L-フェニルアラニンに対するオキシダーゼ活性」及び「L-トリプトファンに対するオキシダーゼ活性」は、実施例における「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法においてタンパク質構成アミノ酸としてL-フェニルアラニン及びL-トリプトファンをそれぞれ用いた場合に得られる相対活性を意味する。上記「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法で得られる活性値は、何れも検出限界が0.5%である。また、本発明で用いるL-トリプトファンオキシダーゼは、分析精度が高まるという観点から、L-フェニルアラニンに対するオキシダーゼ活性がL-トリプトファンに対するオキシダーゼ活性の0〜2%の範囲であることが好ましく、0〜1.5%の範囲であることがより好ましく、0〜1%の範囲であることがさらに好ましい。
【0021】
さらに「L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない」における「L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対するオキシダーゼ活性」は、実施例における「4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性」の試験方法においてタンパク質構成アミノ酸としてL-トリプトファン及びL-フェニルアラニン以外のアミノ酸をそれぞれ用いた場合に得られる相対活性を意味する。さらに、「オキシダーゼ活性を有さない」は、相対活性が、1%以下であること、好ましくは検出限界(0.5%)を超えた相対活性を示さないことを意味する。
【0022】
尚、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有すること、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さないことは、それぞれ実施例に記載の分析方法(定量方法)を用いることにより確認できる。また、タンパク質構成アミノ酸とは、L-チロシン、L-アラニン、L-システイン、L-アスパラギン酸、L-グルタミン酸、L-グリシン、L-ヒスチジン、L-イソロイシン、L-リジン、L-ロイシン、L-メチオニン、L-アスパラギン、L-プロリン、L-グルタミン、L-アルギニン、L-セリン、L-スレオニン、L-バリン、L-を意味する。
【0023】
本明細書で言う「1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から50個」の範囲は、欠失等を有するタンパク質が、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素であることを意味する。前記「1から50個」の範囲は、前記オキシダーゼ活性を有するタンパク質である割合が高いという観点から、例えば、1から40個、好ましくは1から30個、より好ましくは1から20個、さらに好ましくは1から10個、一層好ましくは1から7個、さらに一層好ましくは1から5個、特に好ましくは1から3個程度であることができる。
【0024】
本明細書で言う「配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列」における同一性は、前記アミノ酸配列の同一性を有するタンパク質が、酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のアミノ酸に対してはオキシダーゼ活性を有さない酵素である限り、特に限定されない。前記アミノ酸配列の同一性は、90%以上であれば特に限定されないが、好ましくは95%以上、さらに好ましくは96%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0025】
本発明で用いるL-トリプトファンオキシダーゼは、特定の種より生産されるものに限らず、特異的にL-トリプトファンのアミノ基を酸化的脱アミノ化するのに伴い過酸化水素を生産する酵素を意味する。
【0026】
さらに、本発明に用いるL-トリプトファンオキシダーゼは、同様の活性を有するものであれば、自然界より分離された生物に由来するもの、本酵素をコードする遺伝子を大腸菌や他の生物を宿主として発現させて得られる酵素またはタンパク質も含まれる。
【0027】
また、異種発現による生産法としては、例えば、同様の活性を有する生物種より抽出したゲノムDNAから該当する遺伝子をPCRにて増幅しpETもしくはpUCなどに組み込んだプラスミドベクターを構築したのち、BL21、JM109などの宿主菌株に形質転換し、培養する方法が挙げられる。これら以外の公知の方法も適宜用いることができる。
【0028】
本発明で用いるL-トリプトファンオキシダーゼの取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組換え技術により作製した組換えタンパク質でもよい。組換えタンパク質を作製する場合には、後述するように当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、上記L-トリプトファンオキシダーゼを産生することができる。
【0029】
上記L-トリプトファンオキシダーゼは、上記L-トリプトファンオキシダーゼをコードする遺伝子をベクター上に搭載し、このベクターによって宿主細胞を形質転換した後、形質転換させた宿主細胞を培養して培養物中に前記遺伝子がコードするタンパク質を蓄積し、蓄積したタンパク質を収集することを含む、生産方法により調製することができる。
【0030】
上記L-トリプトファンオキシダーゼをコードする遺伝子の取得方法は特に限定されない。本発明のL-トリプトファンオキシダーゼをコードする遺伝子は、例えば、配列番号1若しくは2に記載のアミノ酸配列または配列番号3若しくは4に記載した塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0031】
例えば、配列表の配列番号3または4に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、公知の方法を用いて行うことができる。
【0032】
本明細書中の配列表の配列番号1若しくは2に記載したアミノ酸配列または配列番号3若しくは4に示す塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いてクロモバクテリウム・ビオラセウム(Chromobacterium violaceum)NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノムライブラリーをスクリーニングすることにより本発明の遺伝子を単離することができる。ゲノムライブラリーは、クロモバクテリウム・ビオラセウム(Chromobacterium violaceum) NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724から常法により作製することができる。
【0033】
PCR法により本発明のL-トリプトファンオキシダーゼをコードする遺伝子を取得することもできる。上記クロモバクテリウム・ビオラセウム(Chromobacterium violaceum) NBRC 12614またはストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノムライブラリーを鋳型として使用し、配列番号3若しくは4に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌(E. coli)等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0034】
上記したプローブ又はプライマーの調製、ゲノムライブラリーの構築、ゲノムライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知の方法に準じて適宜行うことができる。
【0035】
上記L-トリプトファンオキシダーゼの遺伝子は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製するベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、ベクターは発現ベクターである。発現ベクターにおいて上記遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモータは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0036】
細菌細胞で作動可能なプロモータとしては、バチルス・ステアロテルモフィルス・マルトジェニック・アミラーゼ遺伝子(Geobacillus stearothermophilus maltogenic amylase gene)、バチルス・リケニホルミスαアミラーゼ遺伝子(Bacillus licheniformis alpha-amylase gene)、バチルス・アミロリケファチエンス・BANアミラーゼ遺伝子(Bacillus amyloliquefaciens BAN amylase gene)、バチルス・サブチリス・アルカリプロテアーゼ遺伝子(Bacillus subtilis alkaline protease gene)もしくはバチルス・プミルス・キシロシダーゼ遺伝子(Bacillus pumilus xylosidase gene)のプロモータ、またはファージ・ラムダのPR若しくはPLプロモータ、大腸菌(E. coli)のlac、trp若しくはtacプロモータなどが挙げられる。
【0037】
哺乳動物細胞で作動可能なプロモータの例としては、SV40プロモータ、MT−1(メタロチオネイン遺伝子)プロモータ、またはアデノウイルス2主後期プロモータなどがある。昆虫細胞で作動可能なプロモータの例としては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュウロウイルス即時型初期遺伝子1プロモータ、またはバキュウロウイルス39K遅延型初期遺伝子プロモータ等がある。酵母宿主細胞で作動可能なプロモータの例としては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータなどが挙げられる。糸状菌細胞で作動可能なプロモータの例としては、ADH3プロモータまたはtpiAプロモータなどがある。
【0038】
また、上記L-トリプトファンオキシダーゼの遺伝子は必要に応じて、適切なターミネータに機能的に結合されてもよい。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターは更に、ポリアデニレーションシグナル(例えばSV40またはアデノウイルス5E1b領域由来のもの)、転写エンハンサ配列(例えばSV40エンハンサ)などの要素を有していてもよい。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターは更に、該ベクターが宿主細胞内で複製することを可能にするDNA配列を具備してもよく、その一例としてはSV40複製起点(宿主細胞が哺乳類細胞のとき)が挙げられる。
【0039】
L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、ジヒドロ葉酸レダクターゼ(DHFR)またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、または例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン若しくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。L-トリプトファンオキシダーゼの遺伝子、プロモータ、および所望によりターミネータおよび/または分泌シグナル配列をそれぞれ連結し、これらを適切なベクターに挿入する方法は当業者に周知である。
【0040】
L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。L-トリプトファンオキシダーゼの遺伝子を含む組換えベクターを導入される宿主細胞は、L-トリプトファンオキシダーゼの遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌および高等真核細胞等が挙げられる。
【0041】
細菌細胞の例としては、バチルスまたはストレプトマイセス等のグラム陽性菌又は大腸菌(E. coli)等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、または公知の方法でコンピテント細胞を用いることにより行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も公知であり、例えば、エレクトロポーレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0042】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0043】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0044】
昆虫細胞を宿主として用いる場合には、公知の方法を用いて組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる。
【0045】
バキュロウイルスとしては、例えば、ヨトウガ科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、Spodoptera frugiperdaの卵巣細胞であるSf9、Sf21〔バキュロウイルス・エクスプレッション・ベクターズ、ア・ラボラトリー・マニュアル、ダブリュー・エイチ・フリーマン・アンド・カンパニー(W. H. Freeman and Company)、ニューヨーク(New York)、(1992)〕、Trichoplusia niの卵巣細胞であるHiFive(インビトロジェン社製)等を用いることができる。
組換えウイルスを調製するための、昆虫細胞への組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法又はリポフェクション法等を挙げることができる。
【0046】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明で用いるL-トリプトファンオキシダーゼを単離精製するには、通常のタンパク質の単離、精製法を用いればよい。例えば、本発明で用いるL-トリプトファンオキシダーゼが、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のタンパク質の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、本発明のL-トリプトファンオキシダーゼを精製標品として得ることができる。
【0047】
L-トリプトファンオキシダーゼによるL-トリプトファンの酸化反応を以下の反応式Aに示す。
【化1】
【0048】
本発明で用いるL-トリプトファンオキシダーゼとは、上記式Aに示す反応を触媒する酵素を意味し、従って、その由来やアミノ酸配列の違いで限定されることはない。
【0049】
工程(A)
工程(A)におけるL-トリプトファンオキシダーゼの混合量は、10 mU/ml(トリプトファン1 μmolを1分間で消費する活性を1 Uとする)以上とすることが適当であり、水の混合量は、サンプル中のTrp濃度または反応系中の全質量に応じて適宜決定できるが、例えば、反応系中の全質量の5〜95%の範囲とすることができ、モル比としては、1モルのTrpに対して、例えば、1モル以上の水が存在すればよい。酵素反応を水溶液中で行う場合には過剰量の水が反応系中に存在することになる。L-トリプトファンオキシダーゼの混合量の上限は特にないが、実用的には、例えば、100 mU/ml以下であることができる。しかし、L-トリプトファンオキシダーゼの混合量および水の混合量は、この範囲に限定する意図ではなく、適宜調整できる。
【0050】
さらに、L-トリプトファンオキシダーゼおよび水に加えて、好ましくは、L-トリプトファンオキシダーゼの至適pHを考慮したpHを示す緩衝液を含むことができる。緩衝液を用いる場合には、水は緩衝液として供給することもできる。さらに、L-トリプトファンオキシダーゼの安定化剤をさらに含むこともできる。L-トリプトファンオキシダーゼの安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0051】
工程(B)
工程(B)では、前記混合により得られた反応液を酸素の存在下に所定時間放置する。
L-トリプトファンオキシダーゼによるL-トリプトファン酸化反応においては、反応式Aに示すように、L-トリプトファン脱アミノ化生成物であるインドール-3-ピルビン酸と共に、アンモニア(NH3)と過酸化水素(H2O2)が生成物として得られる。上記反応を、例えば、空気と接触する状態で実施することで、反応液中の溶存酸素として上記酸素は供給される。反応液中酸素を供給する目的で反応液に空気などの酸素含有気体を強制的に供給する必要は通常はない。酵素反応に必要とされる酸素量が微量であり、溶存酸素により十分に賄えるためである。酵素のための放置時間は、例えば、使用する酵素量にもよるが、例えば、10分〜1時間の範囲とすることができる。しかし、この範囲に限定する意図ではなく、適宜調整できる。
【0052】
工程(C)
工程(C)では、放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または反応生成物の少なくとも一種の量を計測する。
存在確認または定量に用いられる生成物が過酸化水素である場合、例えばペルオキシダーゼ反応を用いて測定する方法等の公知の方法により、過酸化水素の存在確認または定量可能である。ペルオキシダーゼ反応を用いて測定する場合、使用可能なペルオキシダーゼは過酸化水素の存在確認または定量に利用可能な酵素であればよく、例えば西洋わさび由来ペルオキシダーゼが挙げられる。また、使用するペルオキシダーゼの基質となり得るものであれば発色剤として使用可能であり、西洋わさび由来ペルオキシダーゼを用いる場合には4-アミノアンチピリン:フェノールなどが挙げられる。西洋わさび由来ペルオキシダーゼを用いる過酸化水素の存在確認または定量のための反応は以下に示す通りである。
【0053】
【化2】
【0054】
4-アミノアンチピリン等の発色剤や蛍光剤は、使用されるペルオキシダーゼの種類によって適宜選択することが可能である。
【0055】
L-トリプトファンオキシダーゼ反応の生成物である過酸化水素は、過酸化水素電極を用いた電流検出型センサーを用いて測定することもできる。この測定により、過酸化水素の存在確認および定量の何れも可能である。過酸化水素電極としては、例えば、ペルオキシダーゼを牛血清アルブミンとともにグルタルアルデヒドに固定化した膜とフェロセンをカーボンペーストに含有させたものを電極として用いるセンサーを挙げることができる。
【0056】
存在確認または定量に用いられる生成物がアンモニアである場合には、アンモニア検出薬を用いて測定することができる。アンモニア検出薬としては、例えば、フェノールと次亜塩素酸の組み合わせによるインドフェノール法を挙げることができる。具体的には、サンプルをフェノール・ニトロプルシド溶液および過塩素酸溶液と混合して発色させることで存在確認ができ、さらに、この発色を635 nmの吸光度を測定することにより、アンモニア定量が可能である。
【0057】
存在確認または定量に用いられる生成物がL-トリプトファンの脱アミノ化生成物であるインドールピルビン酸である場合には、2-オキソ酸還元酵素を用いて、インドールピルビン酸の存在確認または定量を行うことができる。
【0058】
<L-トリプトファンの分析用キット>
本発明は、以下の試薬を含むL-トリプトファンの分析用キットを包含する。
(K1)L-トリプトファンオキシダーゼ
上記L-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。L-トリプトファンの分析には、L-トリプトファンの存在確認および定量を含む。
【0059】
上記L-トリプトファンオキシダーゼは、このL-トリプトファンオキシダーゼの安定化剤との混合物であることもできる。安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。安定化剤は、グリセロールであることが、酵素に対する安定化効果に優れるため好ましい。
【0060】
本発明のキットは、(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5) インドールピルビン酸検出薬の少なくとも一つをさらに含むことができる。
【0061】
(K2)反応用緩衝液は、反応液中を定量等の反応に適したpHに維持するために用いられる。後述の実施例に示すL-トリプトファンオキシダーゼは、至適pHが8〜9の範囲があるので、この範囲のpHを有する緩衝液であることが望ましい。
【0062】
(K3)過酸化水素検出用試薬は、過酸化水素の検出を、例えば、発色もしくは蛍光によって行う場合に用いる。過酸化水素検出用試薬としては、例えば、ペルオキシダーゼとその基質となり得る発色剤の組合せであることができる。具体的には、西洋わさびペルオキシダーゼと2-アミノアンチピリン・フェノールの組み合わせを挙げることができる。
【0063】
(K4)アンモニア検出薬としては、例えば、フェノールと次亜塩素酸の組み合わせによるインドフェノール法を挙げることができる。
【0064】
(K5)インドールピルビン酸検出薬としては、例えば、2-オキソ酸還元酵素を用いることができる。
【0065】
<L-トリプトファンの分析用組成物>
本発明は、L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物も包含する。この組成物に含まれるL-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。L-トリプトファンの分析には、L-トリプトファンの存在確認および定量を含む。本発明の分析用組成物は、前記L-トリプトファンに加えて、例えば、前記酵素の安定化剤、緩衝液または緩衝剤などを含有することができる。
【0066】
このL-トリプトファンオキシダーゼの安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。安定化剤は、グリセロールであることが、酵素に対する安定化効果に優れるため好ましい。
【0067】
酵素と安定化剤との混合比は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲であり、より好ましくは30〜50質量部の範囲である。
【0068】
<酵素センサー>
本発明は、L-トリプトファンオキシダーゼを用いたL-トリプトファンの検出または定量用酵素センサーを包含する。この酵素センサーに用いるL-トリプトファンオキシダーゼは、前記L-トリプトファンの定量方法で説明したL-トリプトファンオキシダーゼと同様のものである。
【0069】
前記検出用電極は過酸化水素検出用電極である。過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極であることができる。L-トリプトファンオキシダーゼがL-トリプトファンと反応することで、過酸化水素が生成するので、この過酸化水素を過酸化水素検出用電極で検出することができる。酵素式過酸化水素電極としては、例えば、ペルオキシダーゼを牛血清アルブミンとともにグルタルアルデヒドに固定化した膜とフェロセンをカーボンペーストに含有させたものを電極として用いるセンサーを挙げることができる。隔膜式過酸化水素電極は、隔膜により過酸化水素と反応する電極が隔離されたタイプの電極である。
【0070】
前記L-トリプトファンオキシダーゼは、L-トリプトファンの検出または定量をより高精度に行うという観点から、検出用電極の表面または検出用電極の近傍に配置されることが好ましく、検出用電極の表面に配置される場合には、検出用電極の表面に固定化されても固定化されなくてもよい。検出用電極の表面に固定化されることで、本発明のセンサーを繰り返し利用できる利点はある。さらに、前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置されることもできる。前記安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【実施例】
【0071】
以下に実施例を挙げて本発明を具体的に説明するが本発明はこれに限定されるものではない。
【0072】
1.L-トリプトファンオキシダーゼの調製例
クロモバクテリウム・ビオラセウム(Chromobacterium violaceum)NBRC 12614ゲノムDNAを調製し、これを鋳型として塩基配列データベース上の各菌の配列(AF172851.1)を元に設計した配列番号5および6のプライマーを用いてPCRを行い、L-トリプトファンオキシダーゼ遺伝子vioAを増幅した。増幅産物をpET-28aに挿入し、vioA発現用プラスミドとした。また、ストレプトマイセス・エスピー(Streptomyces sp.)TA-A0724のゲノム断片を含むコスミドpTYMCsta(非特許文献D)をStuI処理した断片を、HincIIで処理したベクターpTYM19(非特許文献K)に挿入し、L-トリプトファンオキシダーゼ遺伝子staOをサブクローニングした。Stratagene社製QuikChange site-directed mutagenesis kitと配列番号7〜10のプライマーを用い、上記サブクローニングプラスミドの配列を改変した。このプラスミドからNdeIおよびHindIII処理によりstaO遺伝子を切り出し、同制限酵素で処理したpET-26bに挿入し、staO発現用プラスミドとした。なお、各酵素のN末またはC末にHis-tagが付加されるようにプライマー設計を行った。各発現用プラスミドで大腸菌BL21 (DE3)株を形質転換し、VioAもしくはStaO異種発現株とした。プライマーの塩基配列は以下の通りである。
VioA-F: ATTCTAGACATATGAAGCATTCTTCCGATATCTG(配列番号5)
VioA-R: AATAAGCTTCGCGGCGATGCGCTG(配列番号6)
StaO-N-sense: TACTGGAGGAAACATATGACGGCACCC(配列番号7)
StaO-N-anti: CAAGGGTGCCGTCATATGTTTCCTCCA(配列番号8)
StaO-C-sense: GACCGGTCGGCGAAGCTTTCTTCGACCTG(配列番号9)
StaO-C-anti: GCAGGTCGAAGAAAGCTTCGCCGACCGGT(配列番号10)
【0073】
各異種発現株を37℃の振盪培養でOD600が0.6〜0.8に達するまで培養し、IPTGを終濃度0.5 mMになるように添加することで発現誘導をかけた。発現誘導後の培養は16℃で16時間行い、可溶性画分に目的酵素が得られた。またStaOに関しては、Takara社製シャペロンプラスミドpG-KJE8を共発現させると収量がさらに向上した。
【0074】
各発現株破砕液上清をGEヘルスケア社製Ni Sepharoseカラムにロードし、20 mM Tris-HCl, 50 mM imidazole (pH 8.0)溶液で洗浄後、20 mM Tris-HCl, 500 mM imidazole (pH 8.0)溶液で溶出させることによって目的酵素を精製・回収した。これらの酵素の塩基配列を決定し、配列番号3および4に示す。さらに酵素のアミノ酸配列を配列番号1および2に示す。
【0075】
2.L-トリプトファンオキシダーゼの活性測定条件例
20 mM トリス−塩酸バッファー (pH 8.0)、5 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、および任意の量のL-トリプトファンオキシダーゼ酵素液を含む反応液を調製し、30℃での505 nmの吸光度変化を測定する。キノンイミン色素のモル吸光係数は6.4 mM-1・cm-1とする。1分間に1 μmolの過酸化水素を生成する活性を1 Uと定義する。本法の検出限界は0.5%である。
【0076】
3.活性のpH依存性
リン酸カリウムバッファー (pH 6.5, 7.0, 7.5, 8.0, 8.5)およびトリス−塩酸バッファー (pH 8.0, 8.5, 9.0)をバッファーとして用い、各バッファー使用時のStaOの活性測定を行った。20 mMの上記のうちいずれかのバッファー、5 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、およびStaO酵素液を含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。その結果、StaOの酵素活性はpH8.0-9.0で最大になることが示された。尚、VioAの至適pHは非特許文献Cに9.25であることが報告されている。
【0077】
4.L-トリプトファンオキシダーゼのアミノ酸に対する基質特異性
20 mM トリス−塩酸バッファー (pH 8.0)、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、0.2〜20 mU/ml L-トリプトファンオキシダーゼと0.5 mMのアミノ酸(20種類のタンパク構成アミノ酸のうちいずれか1種)を含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。得られた測定値から各アミノ酸との相対活性を算出し、表1のような結果となった。本法の検出限界は0.5%である。
【0078】
StaOはL-トリプトファン以外のアミノ酸とは反応性を示さず(相対活性は検出限界以下であった)、L-フェニルアラニンに対してのみ、L-トリプトファンの0.5%〜1%というごく低い相対活性が検出された。またVioAに関してはL-トリプトファン以外のアミノ酸への反応性は検出されなかった(相対活性は検出限界以下であった)。以上のように、StaOとVioAいずれに関しても、L-トリプトファンに対してきわめて高い基質特異性を有していることが示された。
【0079】
5.L-トリプトファンオキシダーゼのL-トリプトファンに対するKm値
20 mM トリス−塩酸バッファー (pH 8.0)、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、0.5 mU/ml L-トリプトファンオキシダーゼと、0, 6.3, 13, 25, 50, 100, 500μMのL-トリプトファンを含む反応液を調製し、30℃での505 nmの吸光度変化を測定した。
【0080】
各L-トリプトファン濃度で算出されたStaOの活性を図1に示す。この結果から、StaOのL-トリプトファンへのKm値は19μMと算出された。またVioAのKm値に関しては、30μMであると報告されている(非特許文献C)。これらの値は、Coprinus sp.由来のL-アミノ酸オキシダーゼの値(650μM)と比べてきわめて低く、また一般的にアミノ酸定量用に用いられている酵素の基質に対するKm値と比べても同等以下に低い値である。このことは、StaOおよびVioAはL-トリプトファン濃度の低いサンプルにおいても定量用酵素として優れた性質を有することを示している。
【0081】
【表1】
【0082】
6.L-トリプトファンの検量線の作成
40 mM トリス−塩酸バッファー (pH 8.0)、2 mMフェノール、2 mM 4-アミノアンチピリン、30 U/ml西洋わさび由来ペルオキシダーゼ、20 mU/ml L-トリプトファンオキシダーゼ酵素液を含む反応液を調製した。これと等量の0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液を標準試料として混合した。30℃で反応を進行させ、30℃での505 nmの吸光度の経時的変化を測定し、反応の進行をモニタリングした。
【0083】
上述のように作製したサンプルの測定結果を図2〜5に示す。StaO, VioAいずれを用いた場合でも、遅くとも反応開始20分後以降はL-トリプトファン濃度と吸光度の間に良好な直線関係が見られ、本反応系で検量線が作成可能であることが示された。また特にStaOサンプルに関しては、反応時間を過剰に長くした場合でも吸光度上昇はほぼ止まるという変化を示した。こうした性質は、たとえ反応時間がばらついたとしても誤差の出にくい、定量用酵素として適した性質と言える。
【0084】
7.ヒト血漿サンプル中のL-トリプトファン定量
7−1.ヒト血漿試料の前処理
ヒト血漿試料はコージンバイオより購入し、−20℃で保管していた3検体を用いた。それぞれ使用直前に解凍し、そのまま、もしくはMicrocon YM-10で限外濾過による除タンパク処理後に、下記のL-トリプトファン定量に供した。
【0085】
7−2.機器分析によるヒト血漿試料中L-トリプトファンの定量
酵素法による定量の比較対象として、超高速アミノ酸分析システムを用いたプレカラム誘導体化法による定量を行った。前述のようにヒト血漿試料を除タンパク質処理したものを超高速アミノ酸分析システムWaters UPLC Amino Acid Analysis Solution systemに供した。サンプルの誘導体化、分析操作などは上記システムの説明書の手順に従った。0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液を標準試料として検量線を作製し、血漿試料中L-トリプトファンの定量を行った。
【0086】
7−3.L-トリプトファンオキシダーゼによるヒト血漿試料中L-トリプトファンの定量
40 mM トリス−塩酸バッファー (pH 8.0)、2 mMフェノール、2 mM 4-アミノアンチピリン、30 U/ml西洋わさび由来ペルオキシダーゼ、20 mU/ml L-トリプトファンオキシダーゼ酵素液を含む反応液を調製した。これと等量の0, 20, 40, 60, 80, 100μMのL-トリプトファン水溶液、もしくは除タンパク処理前後のヒト血漿試料を混合した。30℃で反応を進行させ、30分後に505 nmの吸光度を測定した。L-トリプトファン水溶液サンプルの測定値から検量線を作製し、血漿試料中L-トリプトファンの定量を行った。
【0087】
7−4.機器分析もしくはL-トリプトファンオキシダーゼによるヒト血漿試料の定量結果の比較
上記の測定結果を図6に示す。StaO, VioAいずれの酵素を使った定量系でも、機器分析による測定値に近い値が得られた。よってL-トリプトファンオキシダーゼによる定量法がヒト血漿試料においても有効であることが裏付けられた。
【0088】
一般的な酵素法による生体試料の分析では、あらかじめ除タンパク質処理を行わないと定量性が損なわれるという例が多い。この場合、生体試料の前処理として除タンパク処理が必要となる。一方StaO, VioAはいずれも、除タンパク質処理未処理の試料でも定量性に影響は見られなかった。よって、上記酵素による定量法には試料の前処理が不要であり、酵素法としても特に簡便かつ正確な定量法であることがわかった。
【0089】
8.安定性
StaO, VioA精製酵素液に終濃度0〜50% (v/v)となるようにグリセロールを添加した。各酵素液を4℃で保存し、経時的にサンプリングして活性測定を行った。各種グリセロール濃度存在下におけるStaOまたはVioAの残存活性の経時的変化を図7と図8に示す。StaOに関しては、バッファーのみを含む溶液中では不安定ではあるが、30%以上のグリセロール添加条件で高い安定性を示した。VioAはグリセロール非添加でも比較的安定であるが、10%以上のグリセロールを添加するとその安定性はさらに高まった。
【0090】
VioA精製酵素液に終濃度0または10% (v/v)となるようにグリセロールを添加した。各酵素液を4℃または-80℃で一晩保存した後、活性測定を行った。10%グリセロール存在下または非存在下におけるVioA酵素液の残存活性を図9に示す。VioAの凍結融解時には失活が見られたが、10% グリセロールの添加によりそのような失活は見られなくなった。
【0091】
以上のように、StaOはグリセロール 30%添加、VioAは非添加(凍結融解を行うときには10%添加)で安定な酵素液として使用可能であることが示された。
【0092】
9.反応温度依存性
反応温度20, 30, 40, 50, 60℃でのStaO, VioAの活性測定を行った。20 mM トリス−塩酸バッファー (pH 8.0)、1 mM L-トリプトファン、1 mMフェノール、1 mM 4-アミノアンチピリン、15 U/ml西洋わさび由来ペルオキシダーゼ、およびL-トリプトファンオキシダーゼ酵素液を組成とする活性測定反応液を用いた。上記反応液のうちペルオキシダーゼとL-トリプトファンオキシダーゼ酵素液以外を混合し、上記各種温度で10分間プレインキュベーションを行った。上記反応液の温度を保ったままペルオキシダーゼとL-トリプトファンオキシダーゼ酵素液を添加して反応を開始させ、速やかに505 nmの吸光度変化を測定した。測定結果から算出した、各温度におけるStaO, VioAの相対活性値を表2に示す。両酵素とも、反応温度の上昇と共に活性の上昇が見られた。なお、StaOでは60℃、VioAでは50℃以上で、高温による失活の影響が顕著となり正確な活性測定が困難になった。
【0093】
【表2】
【0094】
10.熱安定性
20%グリセロールを含んだStaO酵素液、および10%グリセロールを含んだVioA酵素液を30, 40, 50, 60, 70℃で1時間熱処理した後、活性測定を行った。熱処理後の各酵素液の残存活性を表3に示す。いずれの酵素も40℃以上の熱処理では顕著な熱失活が見られ、50℃以上の熱処理では残存活性が検出限界以下となった。
【0095】
【表3】
【産業上の利用可能性】
【0096】
L-トリプトファンは、必須アミノ酸であり、極端に不足した場合、ナイアシン欠乏症状を示す事が風土病ペラグラとして知られている。また、過剰に摂取した場合、好酸球増加や筋肉痛症候群の原因になるか可能性が指摘されている。このため、L-トリプトファンの定量は、食品分析、医薬品・サプリメントの品質管理、過剰症・欠乏症時の血液検査及び酵素センサーとして利用が考えられる。また、代謝経路上、ナイアシンやNADなど生体内で重要な機能を持つ物質の原材料となることから、アミノグラムによる疾病診断「アミノインデックス」においても様々な疾病のバイオマーカーを構成する数値の一つとして活用できる。これらの理由から、食品や生体試料などについて、L-トリプトファン量を測定することは、産業的にも医学的にも重要な技術となると考えられるが、現状では、高速液体クロマトフラフィーをはじめとする非常に高価な機器及び試薬が必要となる方法しか実用化されていない。したがって、本発明は、L-トリプトファン定量用キットや酵素センサーなどの形態で商品化が可能であり、安価かつ簡便なL-トリプトファン定量法として事業化できる。
【特許請求の範囲】
【請求項1】
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの分析方法であって、
前記L-トリプトファンオキシダーゼが、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記方法。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項2】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項1に記載の方法。
【請求項3】
工程(A)において検体との混合に用いられる前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤の存在下で保存されているものである、請求項1または2に記載の方法。
【請求項4】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項3に記載の方法。
【請求項5】
前記工程(C)で確認または計測する反応生成物が過酸化水素である請求項1〜4のいずれかに記載の方法。
【請求項6】
以下の試薬を含むL-トリプトファンの分析用キット。
(K1)L-トリプトファンオキシダーゼ
但し、前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項7】
前記(K1)L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項6に記載のキット。
【請求項8】
前記(K1)L-トリプトファンオキシダーゼは、前記酵素の安定化剤との混合物である、請求項6または7に記載のキット。
【請求項9】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項8に記載のキット。
【請求項10】
(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5)インドールピルビン酸検出薬の少なくとも一つをさらに含む、請求項6〜9のいずれかに記載のキット。
【請求項11】
L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物であって、
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記組成物。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項12】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項11に記載の組成物。
【請求項13】
前記酵素の安定化剤を含有する、請求項11または12に記載の組成物。
【請求項14】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項13に記載の組成物。
【請求項15】
L-トリプトファンオキシダーゼを用いるL-トリプトファンの検出または定量用酵素センサーであって、
前記検出用電極は過酸化水素検出用電極であり、かつ
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記センサー。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項16】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項15に記載の酵素センサー。
【請求項17】
前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に用いられる、請求項15または16に記載の酵素センサー。
【請求項18】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項17に記載の酵素センサー。
【請求項19】
過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極である請求項15〜18のいずれかに記載の酵素センサー。
【請求項1】
検体とL-トリプトファンオキシダーゼと水を混合する工程(A)、
前記混合により得られた反応液を酸素の存在下に所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の存在を確認するか、または前記反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの分析方法であって、
前記L-トリプトファンオキシダーゼが、
(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記方法。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項2】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項1に記載の方法。
【請求項3】
工程(A)において検体との混合に用いられる前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤の存在下で保存されているものである、請求項1または2に記載の方法。
【請求項4】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項3に記載の方法。
【請求項5】
前記工程(C)で確認または計測する反応生成物が過酸化水素である請求項1〜4のいずれかに記載の方法。
【請求項6】
以下の試薬を含むL-トリプトファンの分析用キット。
(K1)L-トリプトファンオキシダーゼ
但し、前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項7】
前記(K1)L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項6に記載のキット。
【請求項8】
前記(K1)L-トリプトファンオキシダーゼは、前記酵素の安定化剤との混合物である、請求項6または7に記載のキット。
【請求項9】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項8に記載のキット。
【請求項10】
(K2)反応用緩衝液、(K3)過酸化水素検出用試薬、(K4)アンモニア検出薬および(K5)インドールピルビン酸検出薬の少なくとも一つをさらに含む、請求項6〜9のいずれかに記載のキット。
【請求項11】
L-トリプトファンオキシダーゼを含むL-トリプトファンの分析用組成物であって、
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記組成物。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項12】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項11に記載の組成物。
【請求項13】
前記酵素の安定化剤を含有する、請求項11または12に記載の組成物。
【請求項14】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項13に記載の組成物。
【請求項15】
L-トリプトファンオキシダーゼを用いるL-トリプトファンの検出または定量用酵素センサーであって、
前記検出用電極は過酸化水素検出用電極であり、かつ
前記L-トリプトファンオキシダーゼは、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a2)酸素及び水の存在下、L-トリプトファンに作用して過酸化水素とアンモニアを生成するオキシダーゼ活性を有し、L-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜3%の範囲であり、L-トリプトファン及びL-フェニルアラニン以外のタンパク質構成アミノ酸に対してはオキシダーゼ活性を有さない、前記センサー。
(1)配列表の配列番号1または2に記載のアミノ酸配列;
(2)配列表の配列番号1または2に記載のアミノ酸配列において1から50個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1または2に記載のアミノ酸配列に対して90%以上の同一性を有するアミノ酸配列
【請求項16】
前記L-トリプトファンオキシダーゼの(a1)におけるL-フェニルアラニンに対するオキシダーゼ活性はL-トリプトファンに対するオキシダーゼ活性の0〜1%の範囲である、請求項15に記載の酵素センサー。
【請求項17】
前記L-トリプトファンオキシダーゼは、前記酵素の安定化剤と共に用いられる、請求項15または16に記載の酵素センサー。
【請求項18】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項17に記載の酵素センサー。
【請求項19】
過酸化水素検出用電極は、酵素式過酸化水素電極または隔膜式過酸化水素電極である請求項15〜18のいずれかに記載の酵素センサー。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2012−196207(P2012−196207A)
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願番号】特願2012−46414(P2012−46414)
【出願日】平成24年3月2日(2012.3.2)
【出願人】(000236920)富山県 (197)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願日】平成24年3月2日(2012.3.2)
【出願人】(000236920)富山県 (197)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
[ Back to top ]