
藻類防除剤及び藻類防除方法
【課題】水田や各種プラントにおおける、藻類の発生による生産物や産業製品の生産性や品質の低下、悪臭の発生といった問題を解決する。
【解決手段】植物型ディフェンシンの機能解析を進めていく過程で植物型ディフェンシンが藻類防除活性を有しているといった知見を得るに至った。新規な藻類防除剤はカラシナ由来のディフェンシンを主成分とする。
【解決手段】植物型ディフェンシンの機能解析を進めていく過程で植物型ディフェンシンが藻類防除活性を有しているといった知見を得るに至った。新規な藻類防除剤はカラシナ由来のディフェンシンを主成分とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、藻類防除活性を有するポリペプチドを主成分とする藻類防除剤及び当該ポリペプチドを使用した藻類防除方法に関する。
【背景技術】
【0002】
農作物生産においては高品質植物の安定生産のために化学合成殺菌剤が多用されているが、環境に対する負荷を少なくするために、化学合成殺菌剤への依存度を軽減することが求められている。高品質植物の安定生産を維持しながら、化学合成殺菌剤への依存度を軽減する科学的な方法の1つとして、化学合成農薬の代替として天然物を利用する方法が考えられる。天然物としては、従来から使用されている微生物由来の抗生物質や、食品の防腐剤として使用されている植物由来の低分子成分等を挙げることができる。しかしながら、前者は抗生物質に耐性を示す微生物の出現とその微生物から他生物への耐性形質の伝播(水平伝播)が懸念されること、また後者は単独使用では高品質植物の安定生産を維持できるだけの殺菌力を有していないことから、これらに代わる環境負荷の小さな安全性の高い天然物由来の抗菌、殺菌性物質が望まれていた。
【0003】
天然物由来の抗菌物質である抗菌タンパク質は、病原菌などに対する防御物資として重要な役割を担っている。抗菌タンパク質は、糸状菌、細菌並びにウイルスなどに対して効果を示すが、その活性強度や活性スペクトラムは多様である。現在までに500種以上の抗菌タンパク質が知られ、その機能性および構造上の特徴は多様であり、アミノ酸配列や二次構造の共通性によって分類される。大きくはαへリックス構造の有無で分類され、現在知られている抗菌タンパク質の多くはαへリックス構造をとる。また、1から4個のジスルフィド結合(SS結合)をとる場合が多いことも抗菌タンパク質の特徴である。さらに、アミノ酸として2から9KDaといった非常に短い構造をとることも抗菌タンパク質に共通している。
【0004】
高等動物は免疫系といった病原菌等に対する非常に高度な防御機構を備えているが、昆虫や植物、とりわけその性質上移動の困難な植物については、外敵からの攻撃に対する抗菌タンパク質の重要性は大きいと考えられる。現在までに、コムギ由来のピュロインドリン、オオムギ由来のチオニン、アブラナ科野菜由来の植物型ディフェンシンをはじめ、数種類の植物由来の抗菌タンパク質が知られており、その抗菌活性の研究がすすめられている。
【0005】
なかでもディフェンシンは、抗菌活性の強度やスペクトラムを特徴付けるアミノ酸構造の違いにより、α型(ヒト)、β型(サル)、サペシンA型(昆虫)、植物型などに分類される。ディフェンシンの多くは、3個のSS結合を持ち、多様な抗細菌、糸状菌活性および抗ウイルス活性を示す。植物型ディフェンシンは、4個のSS結合を持つという構造上の特徴から、他のディフェンシンと区別される。植物型ディフェンシンは1990年に存在が知られたのが最初である(F.J.Colilla et al., FEBS Lett., 270, 191, 1990; E.Mendez et al., Eur. J Biochem., 194, 533, 1990:非特許文献1)。当初はその分子量やアミノ酸配列の特徴から、すでに知られていた抗菌タンパク質チオニンと同一視されたが、その後の詳細な構造解析によってチオニンとは異なるタンパク質であることがわかり(M.Bruix et al., Biochemistry, 32,715, 1993:非特許文献2)、ほ乳類や昆虫のディフェンシンとの構造的な類似性から、1995年に植物型ディフェンシンと名付けられた(F.R.Terras et al., Plant Cell, 7, 573, 1995:非特許文献3)。
【0006】
植物型ディフェンシンは45〜54アミノ酸残基からなり、システインに富む小さい塩基性タンパク質で、多種多様な植物種で存在が認められる。そのアミノ酸構造は多様であるが、システイン残基は高度に保存され、サペシンA型(昆虫)ディフェンシンと同様、システインで安定化されたα-ヘリックスモチーフが存在することが特徴である。植物型ディフェンシンの機能面から注意すべきことは、アミノ酸配列や塩基配列に類似性があっても、すべてが抗菌活性を示すわけではないということである。生物学的活性から(1)病原菌の成長を抑制する、(2)病原菌の成長を遅らせる、(3)抗菌活性はなく、α-アミラーゼを阻害する、の3グループに分類される(F.Fant., J. Mol. Biol. 279, 257, 1998:非特許文献4)。また、ダイコン由来のディフェンシンは今までに4種類が知られており、抗菌活性の強い群(2種類)と弱い群(2種類)に明確に分類できる。ディフェンシンタンパク質の抗菌活性は、同種の病原糸状菌に対しても亜種間で大きく異なることが報告されている。例えば、Rhizoctonia solaniに対するダイコン由来ディフェンシンRs-AFP1の抗菌活性を例にとると、生育を50%阻害する濃度が100ppm(F.R.Terras et al., J Biol Chem., 267,15301, 1992:非特許文献5)とする報告と7ppmとする報告の2つの報文(川田ら, 化学と生物, 43, 4, 2005:非特許文献6)が存在する。
【0007】
アブラナ科の野菜(ダイコン)から初めて抗菌活性のあるディフェンシン型タンパク質が見つかったのは1992年である(非特許文献5)。その後、キャベツ、コマツナ、ノザワナ、ハクサイ、カブ、カラシナ、ワサビダイコンおよびナタネ等からディフェンシン遺伝子が単離され、アミノ酸配列を推定された結果、ディフェンシンは共通した特徴を保持しながら、8種類の野菜でいずれも独自のアミノ酸配列を持つことが明らかとなっている(川田元滋ら, 農業および園芸, 78, 470, 2003:非特許文献7)。植物型ディフェンシンのアミノ酸配列について、従来は特定の箇所のアミノ酸残基が共通していることが特徴であるとされていたが、80種にのぼる植物種でディフェンシン遺伝子の存在が知られている現在、そのような配列の共通性はないと考えられている(B.P.Thomma et al., Planta, 216, 193, 2002:非特許文献8)。
【0008】
【非特許文献1】F.J.Colilla et al., FEBS Lett., 270, 191, 1990; E.Mendez et al., Eur. J Biochem., 194, 533, 1990
【非特許文献2】M.Bruix et al., Biochemistry, 32,715, 1993
【非特許文献3】F.R.Terras et al., Plant Cell, 7, 573, 1995
【非特許文献4】F.Fant., J. Mol. Biol. 279, 257, 1998
【非特許文献5】F.R.Terras et al., J Biol Chem., 267,15301, 1992
【非特許文献6】川田ら, 化学と生物, 43, 4, 2005
【非特許文献7】川田元滋ら, 農業および園芸, 78, 470, 2003
【非特許文献8】B.P.Thomma et al., Planta, 216, 193, 2002
【発明の開示】
【発明が解決しようとする課題】
【0009】
上述のように、植物型ディフェンシンに関して、従来種々の研究がなされてきたが、これらの研究の成果としては特定の病原糸状菌を対象とした抗菌活性が知られるのみであった。一方、藻類は水田に発生して表土剥離の原因となり、各種製造プラントに生じてスライムの原因となる。このため、水田や各種プラントにおいては、藻類の発生による生産物や産業製品の生産性や品質の低下、悪臭の発生といった問題が生じていた。特に、植物型ディフェンシンを藻類の防除に利用するといった、植物型ディフェンシンの機能について全く示唆されていなかった。
【0010】
そこで、本発明は、上述したような実情に鑑み、新規な藻類防除剤及び藻類防除方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、植物型ディフェンシンの機能解析を進めていく過程で植物型ディフェンシンが藻類防除活性を有しているといった知見を得る至り、本発明を完成した。本発明は以下を包含する。
(1)カラシナ由来のディフェンシンを主成分とする藻類防除剤。
(2)植物病原糸状菌に対する防除活性を有することを特徴とする(1)記載の藻類防除剤。
(3)上記植物病糸状菌は以下の群から選ばれる少なくとも1種であることを特徴とする(2)記載の藻類防除剤。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
(4)藻類防除対象に対して、カラシナ由来のディフェンシンを主成分とする藻類防除剤を添加する、藻類防除方法。
(5)植物病糸状菌を更に防除することを特徴とする(4)記載の藻類防除方法。
(6)上記植物病糸状菌は以下の群から選ばれる少なくとも1種であることを特徴とする(5)記載の藻類防除方法。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
(7)4〜20ppmとなるように水環境に添加することを特徴とする(4)記載の藻類防除方法。
(8)上記藻類防除対象は、環境及び製造プラントであることを特徴とする(4)記載の藻類防除方法。
【発明の効果】
【0012】
本発明によれば、カラシナ由来のディフェンシンを主成分とする新規な藻類防除剤、及びカラシナ由来のディフェンシンを使用した新規な藻類防除方法を提供することができる。本発明に係る藻類防除剤及び藻類防除方法では、カラシナ由来のディフェンシンといった天然物質を使用しているため、環境負荷の小さく、かつ高い安全性を確保することができる。
【発明を実施するための最良の形態】
【0013】
以下、本発明を詳細に説明する。
本発明に係る藻類防除剤は、カラシナ由来のディフェンシンを主成分とするものである。カラシナ由来のディフェンシンは、カラシナから単離したディフェンシン遺伝子から生産することができる。カラシナからのディフェンシン遺伝子の単離方法の一例を以下に示すが、本発明の技術的範囲は以下の例に限定されるのもではない。すなわち、以下に示す方法以外の方法でディフェンシン遺伝子を単離しても良いことは勿論である。
【0014】
ディフェンシン遺伝子は、例えばカラシナ種子から抽出したRNAから精製したmRNAを用いて、RT-PCR若しくはcDNAライブラリーからのスクリーニングをすることにより得ることができる。但し、ディフェンシン遺伝子の供給源は、カラシナの種子に限定されるものではなく、例えば成長した植物体、植物器官(例えば葉、花弁、茎等)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織等)、又は植物培養細胞(例えばカルス)などであってもよい。さらにディフェンシン遺伝子の供給源とする植物は、カラシナ以外の植物、例えば、限定するものではないが、アブラナ科に属する他の植物等であってもよい。上記植物としては、カラシナが好ましく、mRNAの抽出及びcDNAライブラリーの作製は常法に従って行うことができる。
【0015】
また、mRNAの調製は、通常行われる手法により行うことができる。例えば、上記供給源から、グアニジウムチオシアネート-トリフルオロ酢酸セシウム法などにより全RNAを抽出した後、オリゴdT-セルロースやポリU-セファロース等を用いたアフィニティーカラム法により、あるいはバッチ法によりポリ(A)+RNA(mRNA)を得ることができる。さらに、ショ糖密度勾配遠心法等によりポリ(A)+RNAをさらに分画してもよい。このようにして得られたmRNAを鋳型として、オリゴdTプライマー及び逆転写酵素を用いて一本鎖cDNAを合成した後、該一本鎖cDNAから二本鎖cDNAを合成する。次に、得られた二本鎖cDNAを適当なクローニングベクターに組み込んで組換えベクターを作製する。ベクターにディフェンシン遺伝子のcDNAを挿入するには、まず、精製されたcDNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。そしてこの組換えベクターを用いて大腸菌等を形質転換し、テトラサイクリン耐性、アンピシリン耐性等を指標として形質転換体を選択することにより、cDNAのライブラリーを得ることができる。
【0016】
ここで、大腸菌の形質転換は、Hanahanの方法(J. Mol. Biol. 166:557-580, 1983)、すなわち塩化カルシウム、塩化マグネシウム又は塩化ルビジウムを共存させて調製したコンピテント細胞に、組換えベクターを加える方法等により行うことができる。なお、ベクターとしてプラスミドを用いる場合はテトラサイクリン、アンピシリン等の薬剤耐性遺伝子を含有することが必要である。また、プラスミド以外のクローニングベクター、例えばλファージ(λgt11等)を用いることもできる。
【0017】
プラスミドDNAとしては、大腸菌由来のプラスミド(例えばpBR322、pBR325、pUC118、pUC119、pUC18、pUC19等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YEp24、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP等)が挙げられる。さらに、レトロウイルス又はワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。
【0018】
また、目的のcDNAを有する株を選択するには、例えば、ディフェンシンのアミノ酸配列に対応する縮重センスプライマー及び縮重アンチセンスプライマーを合成し、これを用いてPCRを行う方法も採用することができる。プライマーは化学合成により調製することができる。得られたDNA増幅断片を、32P、35S又はビオチン等で標識してプローブとし、これを形質転換体のDNAを変性固定したニトロセルロースフィルターとハイブリダイズさせ、得られたポジティブ株を検索することによりスクリーニングすることができる。好ましくは、クローニングした部分配列を有するオリゴヌクレオチドと、ライブラリー作製時にcDNAをλファージDNAにライゲーションしたcDNA集団のλファージDNAの配列を有するオリゴヌクレオチドとの間でPCR法を行い、より長い部分配列を取得する。その取得した配列を参考としPCR法によりプローブを作製し、cDNAライブラリーから該遺伝子をスクリーニングすることが望ましい。
【0019】
次に、得られたクローンから全長のcDNAをクローニングする。cDNAのクローニングには、例えばRACE(Rapid Amplification of cDNA ends)法が用いられる。RACE法とは、cDNAの5'又は3'欠失部位をPCRにより迅速に回収する方法である。なお、RACE法は、市販のキット(MarathonTMcDNA Amplification Kit(Clonetech社))を用いて行うこともできる。本発明においては、上記スクリーニングにおいて得られたcDNAの単離クローンについて、PCR産物をテンプレートにしてcDNAの塩基配列を決定する。塩基配列の決定はマキサム-ギルバートの化学修飾法、又はM13ファージを用いるジデオキシヌクレオチド鎖終結法等の公知手法により行うことができるが、通常は自動塩基配列決定装置(例えばApplied Biosystems社製ABI373シークエンサー等)を用いて配列決定が行われる。
【0020】
以上のようにして、カラシナのディフェンシン遺伝子を単離することができる。単離したカラシナのディフェンシン遺伝子を使用することによって、カラシナ由来のディフェンシンを取得することができる。
【0021】
例えば、前述の形質転換体(宿主細胞)を適当な培地で培養し、その培養物からディフェンシンを採取することによって得ることができる。形質転換体の培養は、常法に従って行えばよい。例えば、大腸菌や酵母等の微生物を宿主とする形質転換体の場合は、微生物が資化しうる炭素源、窒素源、無機塩類等を含有し、形質転換体を効率的に培養しうる天然培地、あるいは合成培地で培養すればよい。また、植物細胞を宿主として用いている場合には、チアミン、ピリドキシン等のビタミン類を添加した植物細胞用の培地で培養すればよい。
【0022】
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類が用いられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩またはその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が用いられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等が用いられる。
【0023】
培地中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、Lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル-β-D-チオガラクトピラノシド(IPTG)等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸(IAA)等を培地に添加してもよい。
【0024】
培養は、通常、振盪培養または通気攪拌培養等の好気的条件下、30〜37℃位で6時間〜3日間程度行う。培養期間中、pHは7.0〜7.5程度に保持する。pHの調整は、無機または有機酸、アルカリ溶液等を用いて行う。培養後、ディフェンシンが菌体内または細胞内に生産される場合には、菌体または細胞を破砕することにより該タンパク質を抽出する。また、ディフェンシンが菌体外または細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体または細胞を除去する。その後、ディフェンシンの単離精製に用いられる一般的な生化学的方法、例えば、硫酸アンモニウム沈殿、SDS-PAGE、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等を単独でまたは適宜組み合わせて用いることにより、前記培養物中からディフェンシンを単離精製することができる。
【0025】
前述のように単離精製したディフェンシンの藻類防除活性は、マイクロプレートリーダーを用いたin vitro系でその活性強度を定量化することができる。
【0026】
一方、本発明に係る藻類防除剤は、前述の単離精製したディフェンシン以外の成分を含むものであってもよい。すなわち、単離精製したディフェンシンを藻類防除剤の有効成分として用いる際には、本タンパク質以外の他成分を何ら加えずに藻類防除剤としてもよく、或いは、本タンパク質以外の他の成分を含む組成物として藻類防除剤を調製してもよい。他の成分としては、前述の形質転換体からディフェンシンを単離精製した場合には当該形質転換体の培養物もしくはその処理物を挙げることができる。ここで培養物とは、例えば形質転換体の培養液、該培養液から遠心分離、ろ過等の分離操作によって分離された菌体または培養上清等を意味し、培養物の処理物とは、該菌体を凍結乾燥機、スプレードライヤー等で乾燥処理した乾燥菌体、前記培養液を限外ろ過や透析などによって培地成分などを一部もしくは全部除去した処理液、菌体の死菌化処理物等をあげることができる。かかる死菌化処理法としては、例えば、物理的殺菌法や、化学薬品を用いる殺菌法を挙げることができ、必要に応じてこれらを適宜組み合わせて用いることができる。
【0027】
さらに、単離精製したディフェンシンを固体担体、液体担体、餌等と混合し、必要であれば界面活性剤、その他の製剤用補助剤(固着剤、分散剤等)を添加することで藻類防除剤としてもよい。藻類防除剤としては、通常採用される製剤形態(例えば、乳剤、水和剤、フロアブル剤、粒剤、粉剤、毒餌等)であって、ディフェンシンの藻類防除性能に適し、ディフェンシンの藻類防除活性が保持され得る製剤形態を選択すればよい。これらの製剤には、有効成分としてディフェンシンを、通常、重量比で0.1%〜99%、好ましくは1%〜90%含有すればよい。
【0028】
製剤化の際に用いられる固体担体としては、たとえば粘土類(カオリンクレー、珪藻土、合成含水酸化珪素、ベントナイト、フバサミクレー、酸性白土等)、タルク類、セラミック、その他の無機鉱物(セリサイト、石英、硫黄、活性炭、炭酸カルシウム、水和シリカ等)、化学肥料(硫安、燐安、硝安、尿素、塩安等)等の微粉末あるいは粒状物等があげられ、液体担体としては、たとえば水、アルコール類、ケトン類、芳香族炭化水素類、エステル類、エーテル類、酸アミド類、ハロゲン化炭化水素類、ジメチルスルホキシド、大豆油、綿実油等の植物油等があげられる。界面活性剤としては、たとえばアルキル硫酸エステル塩、アルキルスルホン酸塩、アルキルアリールスルホン酸塩、アルキルアリールエーテル類およびそのポリオキシエチレン化物、ポリエチレングリコールエーテル類、多価アルコールエステル類、糖アルコール誘導体等があげられる。製剤用補助剤としては、たとえばカゼイン、ゼラチン、多糖類(でんぷん粉、アラビアガム、セルロース誘導体、アルギン酸等)、リグニン誘導体、ベントナイト、糖類、合成水溶性高分子(ポリビニルアルコール、ポリビニルピロリドン、ポリアクリル酸類等)、植物油、鉱物油、脂肪酸またはそのエステル等があげられる。毒餌の基材としては、たとえば穀物粉、植物油、糖、結晶セルロース等の餌成分や、保存料、誤食防止剤、誘引性香料等があげられる。
【0029】
このようにして得られる製剤はそのままであるいは水等に希釈して用いることができる。また、本発明に係る藻類防除剤は、従来公知の他の藻類防除剤、殺菌剤、除草剤、植物生長調節剤、共力剤、肥料、土壌改良剤等と混用または併用することもできる。従来公知の他の藻類防除剤としては、塩素、並びにグルタルアルデヒド、イソチアゾロン系化合物、第4級アンモニウム塩化合物等の有機合成化合物を挙げることができる。
【0030】
また、本発明に係る藻類防除剤により防除効果が期待される藻類としては、限定されないが、クロレラ属やアオミドロ属に代表される緑藻類、アウラコセイラ属に代表される珪藻類を挙げることができる。
【0031】
本発明に係る藻類防除剤の施用手段としては、例えばスプレー、自動噴霧器等による散布、振りかけ、塗り付け、浸漬等が挙げられる。本発明に係る藻類防除剤を、農業用もしくは園芸用として目的の耕作領域(水田を含む)に対して施用する場合、その施用量は、通常、10アールあたりディフェンシン重量で10g〜20gであり、乳剤、水和剤、フロアブル剤等を水で希釈して用いる場合は、その施用濃度は通常、ディフェンシン濃度で100ppm〜200ppmである。なお、これらの施用量、施用濃度は、いずれも製剤の種類、施用時期、施用場所、施用方法、対象とする藻類の種類等の状況によって異なり、上記の範囲に限定されることなく適宜増減させることができる。
【0032】
また、本発明に係る藻類防除剤は、以下に列挙する病原糸状菌に対する抗菌活性を併せ持っている。
【0033】
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【0034】
従って、本発明に係る藻類防除剤は、防除対象となる水環境における前述の病原糸状菌の繁殖を防ぐことができる。
【0035】
また、本発明に係る藻類防除剤は、天然物由来の抗菌物質であるカラシナ由来のディフェンシンを主成分とすることから、環境負荷が小さく安全性に優れたものとして広範囲に利用することができる。例えば藻類は、水田の表土が膜状になって日中水面に浮上する現象(表土剥離)を引き起こしたり、ゴルフ場のグリーンの芝生の生育を弱めたり、製紙パルプ、カゼイン、澱粉糊、にかわ、塗工紙、紙用塗工液、表面サイズ剤、塗料、接着剤、合成ゴムラテックス、インキ、ポリビニルアルコールフィルム、塩化ビニルフィルム、樹脂製品、セメント混和剤、シーリング剤、目地剤などの各種の産業製品製造過程で発生するスライムとなることで、産業製品の生産性や品質の低下、悪臭の発生などの原因となっている。そのため、このような藻類の繁殖を防ぐために、本発明に係る藻類防除剤を使用することができる。
【0036】
よって本発明に係る藻類防除剤は、水田、農地、ゴルフ場、湖沼、池等の水環境や、各種産業製品の製造プラントにおける藻類防除に使用することができる。
【実施例】
【0037】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0038】
<実施例1> ディフェンシン遺伝子の単離及びベクターへのクローニング
カラシナの種子からtotal RNAを抽出し、mRNAを精製した後、RT-PCRで増幅してディフェンシン遺伝子を単離した。さらにこれをプローブとしてcDNAライブラリーのスクリーニングを行い、ディフェンシン遺伝子のcDNA全長を単離した。
【0039】
ディフェンシンcDNAの塩基配列の保存された遺伝子5’領域及びpoly A領域に基づいて合成された1組のプライマー(配列番号1、2)を用いてPCR法によりディフェンシン遺伝子を含む領域を増幅させた。XbaIとSacIを用いた制限酵素処理により突出末端化処理をしたDNA断片を、同酵素を用いて制限酵素処理したpUC19ベクターに連結した。塩基配列の確認の後、本ベクターを更に鋳型DNAとしてBamHIとEcoRI制限酵素認識領域を各々含む2種の合成DNA(配列番号3、4)を用いてPCR法によって増幅断片を取得した。得られた増幅断片を、BamHIとEcoRIを用いた制限酵素処理により突出末端化処理を施し、同酵素を用いて酵素処理をしたGSTタグ融合タンパク質発現ベクターpGEX‐6P‐1(アマシャム)に連結させた。
【0040】
配列番号1:合成DNA
gggtctagaatggcyaagtttgtgtc
配列番号2:合成DNA
cccgagctcttttttttttttttt
配列番号3:合成 DNA
gggggatcccagaagttgtgggagaggcca
(ESP+BAM1:下線部は BamHI 認識領域)
配列番号4:合成 DNA
cccgaattcttaaccaagggaagtagcaaat
(MUT‐17:下線部は EcoRI 認識領域)
【0041】
<実施例2>ディフェンシンタンパク質の調製
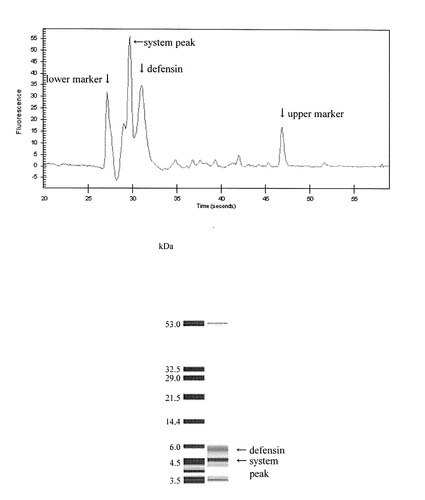
カラシナ由来ディフェンシンタンパク質の活性部位をコードする遺伝子(BJ-AFP1)が組込まれたpGEX-6P-1で形質転換した大腸菌(BL21菌株)を30℃で培養し、600nmの吸光度が0.5〜0.6になったところでIPTGを最終濃度1mMで添加してタンパク質発現を誘導した。その後、2〜3時間培養した後に集菌し、超音波で細胞を破砕し、遠心後上澄みをGSTrap FFカラム(アマシャム)によりアフィニティ精製した。その後、PreScission Protease(アマシャム)でGSTタグを切断し、ディフェンシンを溶出した。この溶出液を陰イオンカラム(Hi-Trap Q FF 1ml(アマシャム))に通し、大腸菌由来のシャペロンタンパク質と考えられるDnaKを除去し、更に塩を取り除くための透析(spectrumフロータ・ライザー, バイオテックCE, MW-cut 2,000)を行った後、遠心濃縮して最終的に蒸留水に溶解した。ディフェンシンの濃度はアジレントバイオアナライザーで測定した。結果を図1に示す。図1に示す結果から本実験により、カラシナ由来のディフェンシンが単離精製できたことが明かとなった。
【0042】
<実施例3> カラシナ由来のディフェンシンの植物病原糸状菌に対する抗菌活性
実施例2で調製したディフェンシン水溶液を用いて下記の被験菌に対する抗菌活性をマイクロプレート法で調べた。具体的には、ポテトデキストロース寒天で7〜10日間培養して着生させた各種糸状菌の胞子懸濁液(1×105〜3×105spore/ml)の10μlを180μlのポテトデキストロース液体培地に添加した後、濃度を変えたディフェンシン水溶液の10μlを添加し、27℃で振とう培養を行った。72時間後に目視或いはマイクロプレートリーダーで650nmの濁度を測定することにより抗菌活性を調べた。
【0043】
供試菌
Alternaria alternata(リンゴ斑点落葉病菌)
Cochliobolus miyabeanus(=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata(=Glomerella rufomaculans-vaccinii、Colletotrichum gloeosporioides)(リンゴ炭疽病菌)
Fulvia fulva(=Cladosporium fulvum)(トマト葉かび病菌)
Pestalotiopsis longisetula(チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae(イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【0044】
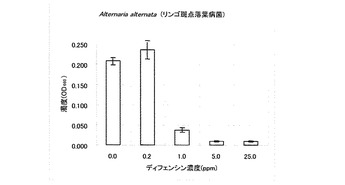
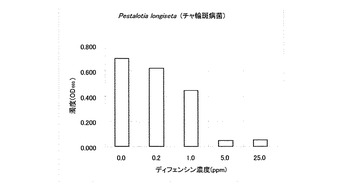
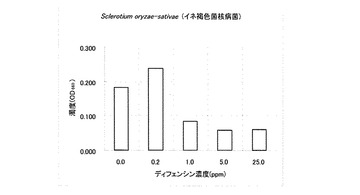
試験結果の一部を表1に示した。T. harzianum、P. debaryanum及びC.miyabeanusの生育は25ppmのディフェンシンにより完全に阻害された。C.miyabeanusの生育は5ppmのディフェンシンでも抑制された。
【0045】
【表1】

【0046】
なお、表1において「++」は無処理と同じくらい生育していることを意味し、「+」は生育抑制が観察されることを意味し、「-」は全く生育が認められないことを意味する。また、その他の供試菌に対する試験結果を図2〜6に示した。A. alternate、P. longiseta、S. oryzae-sativae及びF. fulvaの生育は、1.0ppmのディフェンシンで抑制され、5.0ppmでほぼ完全に阻害された(図2〜5)。また、G. cingulataの生育は0.2ppmで抑制され、1.0ppmでほぼ完全に阻害された(図6)。
【0047】
以上の結果から、カラシナ由来のディフェンシンは、これら病害糸状菌に対する抗菌活性を有することが明かとなった。
【0048】
<実施例4>カラシナ由来のディフェンシンの緑藻類増殖阻害活性
12時間明条件の約20000ルクスの蛍光灯下、28℃でMBM-R液体培地で培養したChlorella sorokiniana(クロレラ)及びScenedesmus acutus(セネデスムス)を集め、液体培地で洗浄後、650nmの吸光度が0.5になるように液体培地で希釈した。一方、様々な濃度でディフェンシンを添加したMBM-R液体培地を96穴マイクロプレートの各ウェルに調製した。緑藻類懸濁液10μlを96穴マイクロプレートの各ウェルに添加して、同じ培養条件で3日間培養した。培養終了後、650nmの濁度を測定するとともに、顕微鏡観察(顕微鏡の倍率:600倍×撮像装置の倍率)を行った。
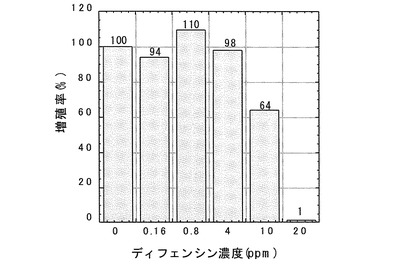
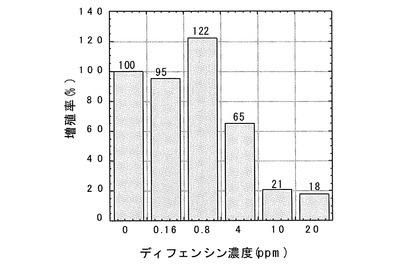
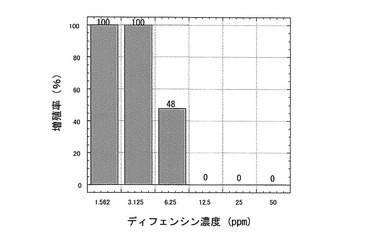
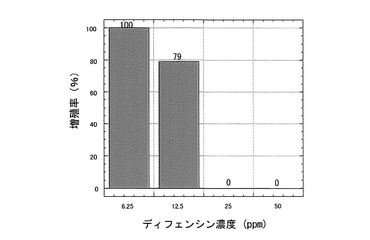
【0049】
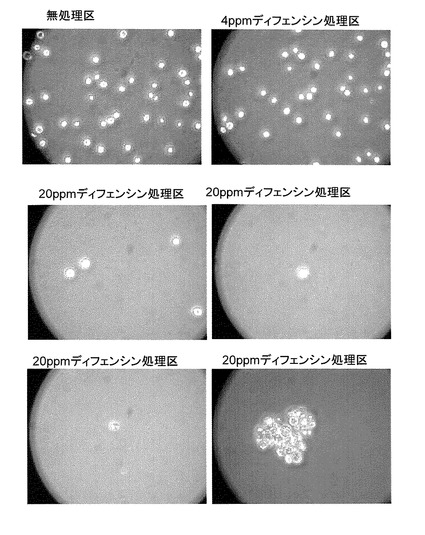
ディフェンシンのクロレラ及びセネデスムスの増殖に対する影響を調べた結果、両者とも20ppmでほぼ完全に阻害された(図7、8)。セネデスムスの増殖は4ppmでも抑制され、クロレラの場合には4ppmでクロロシスが認められた。増殖がほぼ完全に阻害された両緑藻の形態を顕微鏡観察によりディフェンシン無処理区のものと比較したところ、顕著な違いが認められた。増殖が阻害された両緑藻は細胞が膨潤して破裂した後凝集している様子が観察された(図9、10)。この結果は、ディフェンシンが両緑藻の細胞表層(細胞膜或いは細胞壁)に作用していることを示唆した。
【0050】
<実施例5>カラシナ由来のディフェンシンの藍藻類及び珪藻類増殖阻害活性
Merismopedia tenuissima Lemmermann NIES-230(藍藻)及びNitzschia palea (Kutzing) W. Smith(河川から分離した珪藻)を約20000ルクスの蛍光灯の連続照明下23℃で、Csi液体培地中で前培養した。一方、様々な濃度でディフェンシンを添加したCsi液体培地を96穴マイクロプレートの各ウェルに調製した。前培養した藍藻類及び珪藻類懸濁液の10μlを96穴マイクロプレートの各ウェルに添加して、同じ培養条件で3日間培養し、培養終了後に650nmの濁度を測定した。
【0051】
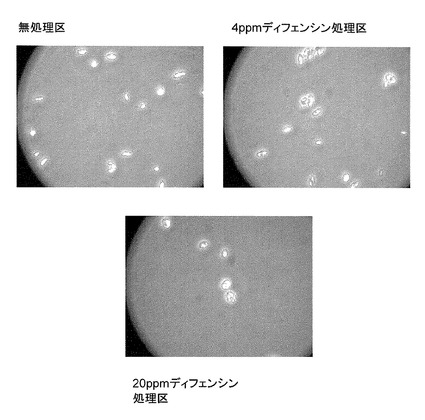
ディフェンシンの藍藻類及び珪藻類の増殖に対する影響を調べた結果、前者は12.5ppmで、後者は25ppmで完全に阻害された(図11、12)
【図面の簡単な説明】
【0052】
【図1】Agilent Bio Analyzer(protein chip 50)によるディフェンシンの泳動パターンであり、上図がエレクトロフェログラムを示し、下図がゲルイメージを示す。
【図2】Alternaria alternata(リンゴ斑点落葉病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図3】Pestalotia longiseta(チャ輪斑病)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図4】Sclerotium oryzae-sativae(イネ褐色菌核病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
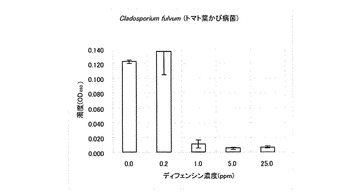
【図5】Fulvia fulva(トマト葉かび病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
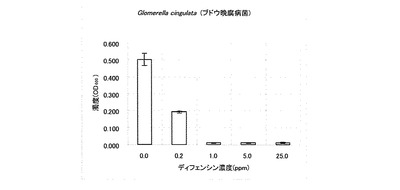
【図6】Glomerella cingulata(ブドウ晩腐病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図7】Chlorella sorokiniana(クロレラ)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図8】Scenedesmus acutus(セネデスムス)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図9】カラシナ由来ディフェンシン処理及び無処理区のChlorella sorokiniana(クロレラ)の顕微鏡写真である。
【図10】カラシナ由来ディフェンシン処理及び無処理区のScenedesmus acutus(セネデスムス)の顕微鏡写真である。
【図11】Merismopedia tenuissima (藍藻)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図12】Nitzschia palea(珪藻)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【技術分野】
【0001】
本発明は、藻類防除活性を有するポリペプチドを主成分とする藻類防除剤及び当該ポリペプチドを使用した藻類防除方法に関する。
【背景技術】
【0002】
農作物生産においては高品質植物の安定生産のために化学合成殺菌剤が多用されているが、環境に対する負荷を少なくするために、化学合成殺菌剤への依存度を軽減することが求められている。高品質植物の安定生産を維持しながら、化学合成殺菌剤への依存度を軽減する科学的な方法の1つとして、化学合成農薬の代替として天然物を利用する方法が考えられる。天然物としては、従来から使用されている微生物由来の抗生物質や、食品の防腐剤として使用されている植物由来の低分子成分等を挙げることができる。しかしながら、前者は抗生物質に耐性を示す微生物の出現とその微生物から他生物への耐性形質の伝播(水平伝播)が懸念されること、また後者は単独使用では高品質植物の安定生産を維持できるだけの殺菌力を有していないことから、これらに代わる環境負荷の小さな安全性の高い天然物由来の抗菌、殺菌性物質が望まれていた。
【0003】
天然物由来の抗菌物質である抗菌タンパク質は、病原菌などに対する防御物資として重要な役割を担っている。抗菌タンパク質は、糸状菌、細菌並びにウイルスなどに対して効果を示すが、その活性強度や活性スペクトラムは多様である。現在までに500種以上の抗菌タンパク質が知られ、その機能性および構造上の特徴は多様であり、アミノ酸配列や二次構造の共通性によって分類される。大きくはαへリックス構造の有無で分類され、現在知られている抗菌タンパク質の多くはαへリックス構造をとる。また、1から4個のジスルフィド結合(SS結合)をとる場合が多いことも抗菌タンパク質の特徴である。さらに、アミノ酸として2から9KDaといった非常に短い構造をとることも抗菌タンパク質に共通している。
【0004】
高等動物は免疫系といった病原菌等に対する非常に高度な防御機構を備えているが、昆虫や植物、とりわけその性質上移動の困難な植物については、外敵からの攻撃に対する抗菌タンパク質の重要性は大きいと考えられる。現在までに、コムギ由来のピュロインドリン、オオムギ由来のチオニン、アブラナ科野菜由来の植物型ディフェンシンをはじめ、数種類の植物由来の抗菌タンパク質が知られており、その抗菌活性の研究がすすめられている。
【0005】
なかでもディフェンシンは、抗菌活性の強度やスペクトラムを特徴付けるアミノ酸構造の違いにより、α型(ヒト)、β型(サル)、サペシンA型(昆虫)、植物型などに分類される。ディフェンシンの多くは、3個のSS結合を持ち、多様な抗細菌、糸状菌活性および抗ウイルス活性を示す。植物型ディフェンシンは、4個のSS結合を持つという構造上の特徴から、他のディフェンシンと区別される。植物型ディフェンシンは1990年に存在が知られたのが最初である(F.J.Colilla et al., FEBS Lett., 270, 191, 1990; E.Mendez et al., Eur. J Biochem., 194, 533, 1990:非特許文献1)。当初はその分子量やアミノ酸配列の特徴から、すでに知られていた抗菌タンパク質チオニンと同一視されたが、その後の詳細な構造解析によってチオニンとは異なるタンパク質であることがわかり(M.Bruix et al., Biochemistry, 32,715, 1993:非特許文献2)、ほ乳類や昆虫のディフェンシンとの構造的な類似性から、1995年に植物型ディフェンシンと名付けられた(F.R.Terras et al., Plant Cell, 7, 573, 1995:非特許文献3)。
【0006】
植物型ディフェンシンは45〜54アミノ酸残基からなり、システインに富む小さい塩基性タンパク質で、多種多様な植物種で存在が認められる。そのアミノ酸構造は多様であるが、システイン残基は高度に保存され、サペシンA型(昆虫)ディフェンシンと同様、システインで安定化されたα-ヘリックスモチーフが存在することが特徴である。植物型ディフェンシンの機能面から注意すべきことは、アミノ酸配列や塩基配列に類似性があっても、すべてが抗菌活性を示すわけではないということである。生物学的活性から(1)病原菌の成長を抑制する、(2)病原菌の成長を遅らせる、(3)抗菌活性はなく、α-アミラーゼを阻害する、の3グループに分類される(F.Fant., J. Mol. Biol. 279, 257, 1998:非特許文献4)。また、ダイコン由来のディフェンシンは今までに4種類が知られており、抗菌活性の強い群(2種類)と弱い群(2種類)に明確に分類できる。ディフェンシンタンパク質の抗菌活性は、同種の病原糸状菌に対しても亜種間で大きく異なることが報告されている。例えば、Rhizoctonia solaniに対するダイコン由来ディフェンシンRs-AFP1の抗菌活性を例にとると、生育を50%阻害する濃度が100ppm(F.R.Terras et al., J Biol Chem., 267,15301, 1992:非特許文献5)とする報告と7ppmとする報告の2つの報文(川田ら, 化学と生物, 43, 4, 2005:非特許文献6)が存在する。
【0007】
アブラナ科の野菜(ダイコン)から初めて抗菌活性のあるディフェンシン型タンパク質が見つかったのは1992年である(非特許文献5)。その後、キャベツ、コマツナ、ノザワナ、ハクサイ、カブ、カラシナ、ワサビダイコンおよびナタネ等からディフェンシン遺伝子が単離され、アミノ酸配列を推定された結果、ディフェンシンは共通した特徴を保持しながら、8種類の野菜でいずれも独自のアミノ酸配列を持つことが明らかとなっている(川田元滋ら, 農業および園芸, 78, 470, 2003:非特許文献7)。植物型ディフェンシンのアミノ酸配列について、従来は特定の箇所のアミノ酸残基が共通していることが特徴であるとされていたが、80種にのぼる植物種でディフェンシン遺伝子の存在が知られている現在、そのような配列の共通性はないと考えられている(B.P.Thomma et al., Planta, 216, 193, 2002:非特許文献8)。
【0008】
【非特許文献1】F.J.Colilla et al., FEBS Lett., 270, 191, 1990; E.Mendez et al., Eur. J Biochem., 194, 533, 1990
【非特許文献2】M.Bruix et al., Biochemistry, 32,715, 1993
【非特許文献3】F.R.Terras et al., Plant Cell, 7, 573, 1995
【非特許文献4】F.Fant., J. Mol. Biol. 279, 257, 1998
【非特許文献5】F.R.Terras et al., J Biol Chem., 267,15301, 1992
【非特許文献6】川田ら, 化学と生物, 43, 4, 2005
【非特許文献7】川田元滋ら, 農業および園芸, 78, 470, 2003
【非特許文献8】B.P.Thomma et al., Planta, 216, 193, 2002
【発明の開示】
【発明が解決しようとする課題】
【0009】
上述のように、植物型ディフェンシンに関して、従来種々の研究がなされてきたが、これらの研究の成果としては特定の病原糸状菌を対象とした抗菌活性が知られるのみであった。一方、藻類は水田に発生して表土剥離の原因となり、各種製造プラントに生じてスライムの原因となる。このため、水田や各種プラントにおいては、藻類の発生による生産物や産業製品の生産性や品質の低下、悪臭の発生といった問題が生じていた。特に、植物型ディフェンシンを藻類の防除に利用するといった、植物型ディフェンシンの機能について全く示唆されていなかった。
【0010】
そこで、本発明は、上述したような実情に鑑み、新規な藻類防除剤及び藻類防除方法を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、植物型ディフェンシンの機能解析を進めていく過程で植物型ディフェンシンが藻類防除活性を有しているといった知見を得る至り、本発明を完成した。本発明は以下を包含する。
(1)カラシナ由来のディフェンシンを主成分とする藻類防除剤。
(2)植物病原糸状菌に対する防除活性を有することを特徴とする(1)記載の藻類防除剤。
(3)上記植物病糸状菌は以下の群から選ばれる少なくとも1種であることを特徴とする(2)記載の藻類防除剤。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
(4)藻類防除対象に対して、カラシナ由来のディフェンシンを主成分とする藻類防除剤を添加する、藻類防除方法。
(5)植物病糸状菌を更に防除することを特徴とする(4)記載の藻類防除方法。
(6)上記植物病糸状菌は以下の群から選ばれる少なくとも1種であることを特徴とする(5)記載の藻類防除方法。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
(7)4〜20ppmとなるように水環境に添加することを特徴とする(4)記載の藻類防除方法。
(8)上記藻類防除対象は、環境及び製造プラントであることを特徴とする(4)記載の藻類防除方法。
【発明の効果】
【0012】
本発明によれば、カラシナ由来のディフェンシンを主成分とする新規な藻類防除剤、及びカラシナ由来のディフェンシンを使用した新規な藻類防除方法を提供することができる。本発明に係る藻類防除剤及び藻類防除方法では、カラシナ由来のディフェンシンといった天然物質を使用しているため、環境負荷の小さく、かつ高い安全性を確保することができる。
【発明を実施するための最良の形態】
【0013】
以下、本発明を詳細に説明する。
本発明に係る藻類防除剤は、カラシナ由来のディフェンシンを主成分とするものである。カラシナ由来のディフェンシンは、カラシナから単離したディフェンシン遺伝子から生産することができる。カラシナからのディフェンシン遺伝子の単離方法の一例を以下に示すが、本発明の技術的範囲は以下の例に限定されるのもではない。すなわち、以下に示す方法以外の方法でディフェンシン遺伝子を単離しても良いことは勿論である。
【0014】
ディフェンシン遺伝子は、例えばカラシナ種子から抽出したRNAから精製したmRNAを用いて、RT-PCR若しくはcDNAライブラリーからのスクリーニングをすることにより得ることができる。但し、ディフェンシン遺伝子の供給源は、カラシナの種子に限定されるものではなく、例えば成長した植物体、植物器官(例えば葉、花弁、茎等)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織等)、又は植物培養細胞(例えばカルス)などであってもよい。さらにディフェンシン遺伝子の供給源とする植物は、カラシナ以外の植物、例えば、限定するものではないが、アブラナ科に属する他の植物等であってもよい。上記植物としては、カラシナが好ましく、mRNAの抽出及びcDNAライブラリーの作製は常法に従って行うことができる。
【0015】
また、mRNAの調製は、通常行われる手法により行うことができる。例えば、上記供給源から、グアニジウムチオシアネート-トリフルオロ酢酸セシウム法などにより全RNAを抽出した後、オリゴdT-セルロースやポリU-セファロース等を用いたアフィニティーカラム法により、あるいはバッチ法によりポリ(A)+RNA(mRNA)を得ることができる。さらに、ショ糖密度勾配遠心法等によりポリ(A)+RNAをさらに分画してもよい。このようにして得られたmRNAを鋳型として、オリゴdTプライマー及び逆転写酵素を用いて一本鎖cDNAを合成した後、該一本鎖cDNAから二本鎖cDNAを合成する。次に、得られた二本鎖cDNAを適当なクローニングベクターに組み込んで組換えベクターを作製する。ベクターにディフェンシン遺伝子のcDNAを挿入するには、まず、精製されたcDNAを適当な制限酵素で切断し、適当なベクターDNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法などが採用される。そしてこの組換えベクターを用いて大腸菌等を形質転換し、テトラサイクリン耐性、アンピシリン耐性等を指標として形質転換体を選択することにより、cDNAのライブラリーを得ることができる。
【0016】
ここで、大腸菌の形質転換は、Hanahanの方法(J. Mol. Biol. 166:557-580, 1983)、すなわち塩化カルシウム、塩化マグネシウム又は塩化ルビジウムを共存させて調製したコンピテント細胞に、組換えベクターを加える方法等により行うことができる。なお、ベクターとしてプラスミドを用いる場合はテトラサイクリン、アンピシリン等の薬剤耐性遺伝子を含有することが必要である。また、プラスミド以外のクローニングベクター、例えばλファージ(λgt11等)を用いることもできる。
【0017】
プラスミドDNAとしては、大腸菌由来のプラスミド(例えばpBR322、pBR325、pUC118、pUC119、pUC18、pUC19等)、枯草菌由来のプラスミド(例えばpUB110、pTP5等)、酵母由来のプラスミド(例えばYEp13、YEp24、YCp50等)などが挙げられ、ファージDNAとしてはλファージ(Charon4A、Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP等)が挙げられる。さらに、レトロウイルス又はワクシニアウイルスなどの動物ウイルス、バキュロウイルスなどの昆虫ウイルスベクターを用いることもできる。
【0018】
また、目的のcDNAを有する株を選択するには、例えば、ディフェンシンのアミノ酸配列に対応する縮重センスプライマー及び縮重アンチセンスプライマーを合成し、これを用いてPCRを行う方法も採用することができる。プライマーは化学合成により調製することができる。得られたDNA増幅断片を、32P、35S又はビオチン等で標識してプローブとし、これを形質転換体のDNAを変性固定したニトロセルロースフィルターとハイブリダイズさせ、得られたポジティブ株を検索することによりスクリーニングすることができる。好ましくは、クローニングした部分配列を有するオリゴヌクレオチドと、ライブラリー作製時にcDNAをλファージDNAにライゲーションしたcDNA集団のλファージDNAの配列を有するオリゴヌクレオチドとの間でPCR法を行い、より長い部分配列を取得する。その取得した配列を参考としPCR法によりプローブを作製し、cDNAライブラリーから該遺伝子をスクリーニングすることが望ましい。
【0019】
次に、得られたクローンから全長のcDNAをクローニングする。cDNAのクローニングには、例えばRACE(Rapid Amplification of cDNA ends)法が用いられる。RACE法とは、cDNAの5'又は3'欠失部位をPCRにより迅速に回収する方法である。なお、RACE法は、市販のキット(MarathonTMcDNA Amplification Kit(Clonetech社))を用いて行うこともできる。本発明においては、上記スクリーニングにおいて得られたcDNAの単離クローンについて、PCR産物をテンプレートにしてcDNAの塩基配列を決定する。塩基配列の決定はマキサム-ギルバートの化学修飾法、又はM13ファージを用いるジデオキシヌクレオチド鎖終結法等の公知手法により行うことができるが、通常は自動塩基配列決定装置(例えばApplied Biosystems社製ABI373シークエンサー等)を用いて配列決定が行われる。
【0020】
以上のようにして、カラシナのディフェンシン遺伝子を単離することができる。単離したカラシナのディフェンシン遺伝子を使用することによって、カラシナ由来のディフェンシンを取得することができる。
【0021】
例えば、前述の形質転換体(宿主細胞)を適当な培地で培養し、その培養物からディフェンシンを採取することによって得ることができる。形質転換体の培養は、常法に従って行えばよい。例えば、大腸菌や酵母等の微生物を宿主とする形質転換体の場合は、微生物が資化しうる炭素源、窒素源、無機塩類等を含有し、形質転換体を効率的に培養しうる天然培地、あるいは合成培地で培養すればよい。また、植物細胞を宿主として用いている場合には、チアミン、ピリドキシン等のビタミン類を添加した植物細胞用の培地で培養すればよい。
【0022】
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類が用いられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩またはその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が用いられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等が用いられる。
【0023】
培地中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、Lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル-β-D-チオガラクトピラノシド(IPTG)等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸(IAA)等を培地に添加してもよい。
【0024】
培養は、通常、振盪培養または通気攪拌培養等の好気的条件下、30〜37℃位で6時間〜3日間程度行う。培養期間中、pHは7.0〜7.5程度に保持する。pHの調整は、無機または有機酸、アルカリ溶液等を用いて行う。培養後、ディフェンシンが菌体内または細胞内に生産される場合には、菌体または細胞を破砕することにより該タンパク質を抽出する。また、ディフェンシンが菌体外または細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体または細胞を除去する。その後、ディフェンシンの単離精製に用いられる一般的な生化学的方法、例えば、硫酸アンモニウム沈殿、SDS-PAGE、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等を単独でまたは適宜組み合わせて用いることにより、前記培養物中からディフェンシンを単離精製することができる。
【0025】
前述のように単離精製したディフェンシンの藻類防除活性は、マイクロプレートリーダーを用いたin vitro系でその活性強度を定量化することができる。
【0026】
一方、本発明に係る藻類防除剤は、前述の単離精製したディフェンシン以外の成分を含むものであってもよい。すなわち、単離精製したディフェンシンを藻類防除剤の有効成分として用いる際には、本タンパク質以外の他成分を何ら加えずに藻類防除剤としてもよく、或いは、本タンパク質以外の他の成分を含む組成物として藻類防除剤を調製してもよい。他の成分としては、前述の形質転換体からディフェンシンを単離精製した場合には当該形質転換体の培養物もしくはその処理物を挙げることができる。ここで培養物とは、例えば形質転換体の培養液、該培養液から遠心分離、ろ過等の分離操作によって分離された菌体または培養上清等を意味し、培養物の処理物とは、該菌体を凍結乾燥機、スプレードライヤー等で乾燥処理した乾燥菌体、前記培養液を限外ろ過や透析などによって培地成分などを一部もしくは全部除去した処理液、菌体の死菌化処理物等をあげることができる。かかる死菌化処理法としては、例えば、物理的殺菌法や、化学薬品を用いる殺菌法を挙げることができ、必要に応じてこれらを適宜組み合わせて用いることができる。
【0027】
さらに、単離精製したディフェンシンを固体担体、液体担体、餌等と混合し、必要であれば界面活性剤、その他の製剤用補助剤(固着剤、分散剤等)を添加することで藻類防除剤としてもよい。藻類防除剤としては、通常採用される製剤形態(例えば、乳剤、水和剤、フロアブル剤、粒剤、粉剤、毒餌等)であって、ディフェンシンの藻類防除性能に適し、ディフェンシンの藻類防除活性が保持され得る製剤形態を選択すればよい。これらの製剤には、有効成分としてディフェンシンを、通常、重量比で0.1%〜99%、好ましくは1%〜90%含有すればよい。
【0028】
製剤化の際に用いられる固体担体としては、たとえば粘土類(カオリンクレー、珪藻土、合成含水酸化珪素、ベントナイト、フバサミクレー、酸性白土等)、タルク類、セラミック、その他の無機鉱物(セリサイト、石英、硫黄、活性炭、炭酸カルシウム、水和シリカ等)、化学肥料(硫安、燐安、硝安、尿素、塩安等)等の微粉末あるいは粒状物等があげられ、液体担体としては、たとえば水、アルコール類、ケトン類、芳香族炭化水素類、エステル類、エーテル類、酸アミド類、ハロゲン化炭化水素類、ジメチルスルホキシド、大豆油、綿実油等の植物油等があげられる。界面活性剤としては、たとえばアルキル硫酸エステル塩、アルキルスルホン酸塩、アルキルアリールスルホン酸塩、アルキルアリールエーテル類およびそのポリオキシエチレン化物、ポリエチレングリコールエーテル類、多価アルコールエステル類、糖アルコール誘導体等があげられる。製剤用補助剤としては、たとえばカゼイン、ゼラチン、多糖類(でんぷん粉、アラビアガム、セルロース誘導体、アルギン酸等)、リグニン誘導体、ベントナイト、糖類、合成水溶性高分子(ポリビニルアルコール、ポリビニルピロリドン、ポリアクリル酸類等)、植物油、鉱物油、脂肪酸またはそのエステル等があげられる。毒餌の基材としては、たとえば穀物粉、植物油、糖、結晶セルロース等の餌成分や、保存料、誤食防止剤、誘引性香料等があげられる。
【0029】
このようにして得られる製剤はそのままであるいは水等に希釈して用いることができる。また、本発明に係る藻類防除剤は、従来公知の他の藻類防除剤、殺菌剤、除草剤、植物生長調節剤、共力剤、肥料、土壌改良剤等と混用または併用することもできる。従来公知の他の藻類防除剤としては、塩素、並びにグルタルアルデヒド、イソチアゾロン系化合物、第4級アンモニウム塩化合物等の有機合成化合物を挙げることができる。
【0030】
また、本発明に係る藻類防除剤により防除効果が期待される藻類としては、限定されないが、クロレラ属やアオミドロ属に代表される緑藻類、アウラコセイラ属に代表される珪藻類を挙げることができる。
【0031】
本発明に係る藻類防除剤の施用手段としては、例えばスプレー、自動噴霧器等による散布、振りかけ、塗り付け、浸漬等が挙げられる。本発明に係る藻類防除剤を、農業用もしくは園芸用として目的の耕作領域(水田を含む)に対して施用する場合、その施用量は、通常、10アールあたりディフェンシン重量で10g〜20gであり、乳剤、水和剤、フロアブル剤等を水で希釈して用いる場合は、その施用濃度は通常、ディフェンシン濃度で100ppm〜200ppmである。なお、これらの施用量、施用濃度は、いずれも製剤の種類、施用時期、施用場所、施用方法、対象とする藻類の種類等の状況によって異なり、上記の範囲に限定されることなく適宜増減させることができる。
【0032】
また、本発明に係る藻類防除剤は、以下に列挙する病原糸状菌に対する抗菌活性を併せ持っている。
【0033】
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【0034】
従って、本発明に係る藻類防除剤は、防除対象となる水環境における前述の病原糸状菌の繁殖を防ぐことができる。
【0035】
また、本発明に係る藻類防除剤は、天然物由来の抗菌物質であるカラシナ由来のディフェンシンを主成分とすることから、環境負荷が小さく安全性に優れたものとして広範囲に利用することができる。例えば藻類は、水田の表土が膜状になって日中水面に浮上する現象(表土剥離)を引き起こしたり、ゴルフ場のグリーンの芝生の生育を弱めたり、製紙パルプ、カゼイン、澱粉糊、にかわ、塗工紙、紙用塗工液、表面サイズ剤、塗料、接着剤、合成ゴムラテックス、インキ、ポリビニルアルコールフィルム、塩化ビニルフィルム、樹脂製品、セメント混和剤、シーリング剤、目地剤などの各種の産業製品製造過程で発生するスライムとなることで、産業製品の生産性や品質の低下、悪臭の発生などの原因となっている。そのため、このような藻類の繁殖を防ぐために、本発明に係る藻類防除剤を使用することができる。
【0036】
よって本発明に係る藻類防除剤は、水田、農地、ゴルフ場、湖沼、池等の水環境や、各種産業製品の製造プラントにおける藻類防除に使用することができる。
【実施例】
【0037】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0038】
<実施例1> ディフェンシン遺伝子の単離及びベクターへのクローニング
カラシナの種子からtotal RNAを抽出し、mRNAを精製した後、RT-PCRで増幅してディフェンシン遺伝子を単離した。さらにこれをプローブとしてcDNAライブラリーのスクリーニングを行い、ディフェンシン遺伝子のcDNA全長を単離した。
【0039】
ディフェンシンcDNAの塩基配列の保存された遺伝子5’領域及びpoly A領域に基づいて合成された1組のプライマー(配列番号1、2)を用いてPCR法によりディフェンシン遺伝子を含む領域を増幅させた。XbaIとSacIを用いた制限酵素処理により突出末端化処理をしたDNA断片を、同酵素を用いて制限酵素処理したpUC19ベクターに連結した。塩基配列の確認の後、本ベクターを更に鋳型DNAとしてBamHIとEcoRI制限酵素認識領域を各々含む2種の合成DNA(配列番号3、4)を用いてPCR法によって増幅断片を取得した。得られた増幅断片を、BamHIとEcoRIを用いた制限酵素処理により突出末端化処理を施し、同酵素を用いて酵素処理をしたGSTタグ融合タンパク質発現ベクターpGEX‐6P‐1(アマシャム)に連結させた。
【0040】
配列番号1:合成DNA
gggtctagaatggcyaagtttgtgtc
配列番号2:合成DNA
cccgagctcttttttttttttttt
配列番号3:合成 DNA
gggggatcccagaagttgtgggagaggcca
(ESP+BAM1:下線部は BamHI 認識領域)
配列番号4:合成 DNA
cccgaattcttaaccaagggaagtagcaaat
(MUT‐17:下線部は EcoRI 認識領域)
【0041】
<実施例2>ディフェンシンタンパク質の調製
カラシナ由来ディフェンシンタンパク質の活性部位をコードする遺伝子(BJ-AFP1)が組込まれたpGEX-6P-1で形質転換した大腸菌(BL21菌株)を30℃で培養し、600nmの吸光度が0.5〜0.6になったところでIPTGを最終濃度1mMで添加してタンパク質発現を誘導した。その後、2〜3時間培養した後に集菌し、超音波で細胞を破砕し、遠心後上澄みをGSTrap FFカラム(アマシャム)によりアフィニティ精製した。その後、PreScission Protease(アマシャム)でGSTタグを切断し、ディフェンシンを溶出した。この溶出液を陰イオンカラム(Hi-Trap Q FF 1ml(アマシャム))に通し、大腸菌由来のシャペロンタンパク質と考えられるDnaKを除去し、更に塩を取り除くための透析(spectrumフロータ・ライザー, バイオテックCE, MW-cut 2,000)を行った後、遠心濃縮して最終的に蒸留水に溶解した。ディフェンシンの濃度はアジレントバイオアナライザーで測定した。結果を図1に示す。図1に示す結果から本実験により、カラシナ由来のディフェンシンが単離精製できたことが明かとなった。
【0042】
<実施例3> カラシナ由来のディフェンシンの植物病原糸状菌に対する抗菌活性
実施例2で調製したディフェンシン水溶液を用いて下記の被験菌に対する抗菌活性をマイクロプレート法で調べた。具体的には、ポテトデキストロース寒天で7〜10日間培養して着生させた各種糸状菌の胞子懸濁液(1×105〜3×105spore/ml)の10μlを180μlのポテトデキストロース液体培地に添加した後、濃度を変えたディフェンシン水溶液の10μlを添加し、27℃で振とう培養を行った。72時間後に目視或いはマイクロプレートリーダーで650nmの濁度を測定することにより抗菌活性を調べた。
【0043】
供試菌
Alternaria alternata(リンゴ斑点落葉病菌)
Cochliobolus miyabeanus(=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata(=Glomerella rufomaculans-vaccinii、Colletotrichum gloeosporioides)(リンゴ炭疽病菌)
Fulvia fulva(=Cladosporium fulvum)(トマト葉かび病菌)
Pestalotiopsis longisetula(チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae(イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【0044】
試験結果の一部を表1に示した。T. harzianum、P. debaryanum及びC.miyabeanusの生育は25ppmのディフェンシンにより完全に阻害された。C.miyabeanusの生育は5ppmのディフェンシンでも抑制された。
【0045】
【表1】
【0046】
なお、表1において「++」は無処理と同じくらい生育していることを意味し、「+」は生育抑制が観察されることを意味し、「-」は全く生育が認められないことを意味する。また、その他の供試菌に対する試験結果を図2〜6に示した。A. alternate、P. longiseta、S. oryzae-sativae及びF. fulvaの生育は、1.0ppmのディフェンシンで抑制され、5.0ppmでほぼ完全に阻害された(図2〜5)。また、G. cingulataの生育は0.2ppmで抑制され、1.0ppmでほぼ完全に阻害された(図6)。
【0047】
以上の結果から、カラシナ由来のディフェンシンは、これら病害糸状菌に対する抗菌活性を有することが明かとなった。
【0048】
<実施例4>カラシナ由来のディフェンシンの緑藻類増殖阻害活性
12時間明条件の約20000ルクスの蛍光灯下、28℃でMBM-R液体培地で培養したChlorella sorokiniana(クロレラ)及びScenedesmus acutus(セネデスムス)を集め、液体培地で洗浄後、650nmの吸光度が0.5になるように液体培地で希釈した。一方、様々な濃度でディフェンシンを添加したMBM-R液体培地を96穴マイクロプレートの各ウェルに調製した。緑藻類懸濁液10μlを96穴マイクロプレートの各ウェルに添加して、同じ培養条件で3日間培養した。培養終了後、650nmの濁度を測定するとともに、顕微鏡観察(顕微鏡の倍率:600倍×撮像装置の倍率)を行った。
【0049】
ディフェンシンのクロレラ及びセネデスムスの増殖に対する影響を調べた結果、両者とも20ppmでほぼ完全に阻害された(図7、8)。セネデスムスの増殖は4ppmでも抑制され、クロレラの場合には4ppmでクロロシスが認められた。増殖がほぼ完全に阻害された両緑藻の形態を顕微鏡観察によりディフェンシン無処理区のものと比較したところ、顕著な違いが認められた。増殖が阻害された両緑藻は細胞が膨潤して破裂した後凝集している様子が観察された(図9、10)。この結果は、ディフェンシンが両緑藻の細胞表層(細胞膜或いは細胞壁)に作用していることを示唆した。
【0050】
<実施例5>カラシナ由来のディフェンシンの藍藻類及び珪藻類増殖阻害活性
Merismopedia tenuissima Lemmermann NIES-230(藍藻)及びNitzschia palea (Kutzing) W. Smith(河川から分離した珪藻)を約20000ルクスの蛍光灯の連続照明下23℃で、Csi液体培地中で前培養した。一方、様々な濃度でディフェンシンを添加したCsi液体培地を96穴マイクロプレートの各ウェルに調製した。前培養した藍藻類及び珪藻類懸濁液の10μlを96穴マイクロプレートの各ウェルに添加して、同じ培養条件で3日間培養し、培養終了後に650nmの濁度を測定した。
【0051】
ディフェンシンの藍藻類及び珪藻類の増殖に対する影響を調べた結果、前者は12.5ppmで、後者は25ppmで完全に阻害された(図11、12)
【図面の簡単な説明】
【0052】
【図1】Agilent Bio Analyzer(protein chip 50)によるディフェンシンの泳動パターンであり、上図がエレクトロフェログラムを示し、下図がゲルイメージを示す。
【図2】Alternaria alternata(リンゴ斑点落葉病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図3】Pestalotia longiseta(チャ輪斑病)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図4】Sclerotium oryzae-sativae(イネ褐色菌核病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図5】Fulvia fulva(トマト葉かび病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図6】Glomerella cingulata(ブドウ晩腐病菌)に対するカラシナ由来ディフェンシンの抗菌活性を示す特性図である。
【図7】Chlorella sorokiniana(クロレラ)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図8】Scenedesmus acutus(セネデスムス)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図9】カラシナ由来ディフェンシン処理及び無処理区のChlorella sorokiniana(クロレラ)の顕微鏡写真である。
【図10】カラシナ由来ディフェンシン処理及び無処理区のScenedesmus acutus(セネデスムス)の顕微鏡写真である。
【図11】Merismopedia tenuissima (藍藻)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【図12】Nitzschia palea(珪藻)増殖に対するカラシナ由来ディフェンシンの阻害活性を示す特性図である。
【特許請求の範囲】
【請求項1】
カラシナ由来のディフェンシンを主成分とする藻類防除剤。
【請求項2】
植物病原糸状菌に対する防除活性を有することを特徴とする請求項1記載の藻類防除剤。
【請求項3】
上記植物病原糸状菌は以下の群から選ばれる少なくとも一種であることを特徴とする請求項2記載の藻類防除剤。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【請求項4】
藻類防除対象に対して、カラシナ由来のディフェンシンを主成分とする藻類防除剤を添加する、藻類防除方法。
【請求項5】
植物病原糸状菌を更に防除することを特徴とする請求項4記載の藻類防除方法。
【請求項6】
上記植物病糸状菌は以下の群から選ばれる少なくとも一種であることを特徴とする請求項5記載の藻類防除方法。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【請求項7】
4〜20ppmとなるように水環境に添加することを特徴とする請求項4記載の藻類防除方法。
【請求項8】
上記藻類防除対象は、環境及び製造プラントであることを特徴とする請求項4記載の藻類防除方法。
【請求項1】
カラシナ由来のディフェンシンを主成分とする藻類防除剤。
【請求項2】
植物病原糸状菌に対する防除活性を有することを特徴とする請求項1記載の藻類防除剤。
【請求項3】
上記植物病原糸状菌は以下の群から選ばれる少なくとも一種であることを特徴とする請求項2記載の藻類防除剤。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【請求項4】
藻類防除対象に対して、カラシナ由来のディフェンシンを主成分とする藻類防除剤を添加する、藻類防除方法。
【請求項5】
植物病原糸状菌を更に防除することを特徴とする請求項4記載の藻類防除方法。
【請求項6】
上記植物病糸状菌は以下の群から選ばれる少なくとも一種であることを特徴とする請求項5記載の藻類防除方法。
Alternaria alternata (リンゴ斑点落葉病菌)
Cochliobolus miyabeanus (=Helminthosporium oryzae)(イネごま葉枯病菌)
Glomerella cingulata (=Glomerella rufomaculans-vaccinii, Colletotrichum gloeosporioides) (リンゴ炭疽病菌)
Fulvia fulva (=Cladosporium fulvum) (トマト葉かび病菌)
Pestalotiopsis longisetula (チャ輪斑病)
Pythium debaryanum(キュウリ苗立枯病菌)
Sclerotium oryzae-sativae (イネ褐色菌核病菌)
Trichoderma harzianum(トリコデルマ菌)
【請求項7】
4〜20ppmとなるように水環境に添加することを特徴とする請求項4記載の藻類防除方法。
【請求項8】
上記藻類防除対象は、環境及び製造プラントであることを特徴とする請求項4記載の藻類防除方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2008−133203(P2008−133203A)
【公開日】平成20年6月12日(2008.6.12)
【国際特許分類】
【出願番号】特願2006−318851(P2006−318851)
【出願日】平成18年11月27日(2006.11.27)
【出願人】(000000169)クミアイ化学工業株式会社 (86)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【出願人】(390034348)ケイ・アイ化成株式会社 (19)
【Fターム(参考)】
【公開日】平成20年6月12日(2008.6.12)
【国際特許分類】
【出願日】平成18年11月27日(2006.11.27)
【出願人】(000000169)クミアイ化学工業株式会社 (86)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【出願人】(390034348)ケイ・アイ化成株式会社 (19)
【Fターム(参考)】
[ Back to top ]