
金属抽出ペプチド(MAP)タグおよび関連する方法
【課題】画像化、研究、化学療法、およびキレーション療法(chelation therapy)等における種々の用途に有用な、選択的に金属に結合する能力を有する、短いペプチドモチーフおよびこのようなモチーフを使用するための方法を提供する。
【解決手段】特定の実施形態において、本発明は、新規なペプチドベースのタグもしくは金属の結合剤(MAPタグ)を提供する。上記MAPタグは、ポリペプチドもしくはタンパク質中に高親和性金属結合部位を直接コードするために使用され得る。本開示のMAPタグは、平面四角形および/もしくは四角錐の幾何的配置において金属に結合し得るそれらの能力によって特徴付けられる。
【解決手段】特定の実施形態において、本発明は、新規なペプチドベースのタグもしくは金属の結合剤(MAPタグ)を提供する。上記MAPタグは、ポリペプチドもしくはタンパク質中に高親和性金属結合部位を直接コードするために使用され得る。本開示のMAPタグは、平面四角形および/もしくは四角錐の幾何的配置において金属に結合し得るそれらの能力によって特徴付けられる。
【発明の詳細な説明】
【技術分野】
【0001】
(関連する出願との相互参照)
本願は、米国仮特許出願第61/052,918号(2008年5月13日出願)に対する優先権を主張する。この仮特許出願は、参照により本願に援用される。
【0002】
(政府の利益の陳述)
本発明は合衆国国立衛生研究所の助成金(Grant No.P20 RR−17708)の下、開発された。合衆国政府は本発明に対し一定の権利を有し得る。
【背景技術】
【0003】
(背景)
以前に、金属を利用して、ポリペプチド(例えば、タンパク質もしくはタンパク質フラグメント)を組成物から抽出することは、一般的な技術であった。このような抽出物は、上記組成物内の他の物質に優先して、金属が特定のポリペプチドと錯体形成する能力に基づいた。2つの異なるペプチドベースの金属結合タグが、組成物からペプチドを分離するために使用され得ることは、公知である。しかし、これら結合タグは、アミノ酸の特定のタイプ、および本発明に関連して記載されるものとは実質的に異なる結合を達成するために利用されるアミノ酸の特定の数もしくは特定の配列を有する。例えば、例えば、これら2種の結合タグは、同じタイプの金属に結合しなくてもよいし、本発明と同じ幾何的配置(geometry)を使用して結合しなくてもよい。第1の結合タグに関するさらなる情報は、特許文献1(Haroonら)において得られ得、配列NXEQVSPを含むペプチドに関する。第2の結合タグに関する情報は、特許文献2(Arbogastら)において得られ得、そこで記載されるタンパク質全体であるようであるタグの配列に関する。
【0004】
タンパク質の組換え発現のために生物の遺伝物質においてコードされ得るタグは、タンパク質生成物の精製および同定のために広く利用されてきた。この最も顕著な例は、His−タグ技術であった。この技術は、固定化金属アフィニティークロマトグラフィー(IMAC)を使用して、全細胞からタグ化したタンパク質を効率的に単離する容易な手段を提供する。多くの他のペプチドベースのタグは、ペプチドタグを認識する抗体を使用して、細胞培養アッセイもしくは細胞溶解物において上記タグ化タンパク質を検出するために開発されてきた。これら技術は、インサイチュアッセイもしくはインビトロアッセイにおいて有用であるが、一般に、それらはインビボ分析には適用可能でない。ペプチドタグの利点は、上記タグが、目的のタンパク質を標識するためのさらなる化学工程の必要なくして、上記タンパク質に共有結合することである。
【0005】
MRI画像化は、生きている動物およびヒトにおいて構造的特徴を調べるための一般的な方法である。上記技術は安全でありかつ非侵襲性である。造影剤は、上記方法の感度を改善するために開発されてきており、それらは、上記MRI画像において認められる特徴を増強するために使用される。上記改善は、造影剤が、隣り合う分子(代表的には、水)からのシグナルを変化させる金属を含むことから、達成されてきた。最も一般に使用される薬剤は、Gd(III)をキレート化するが、他の金属は、対比を増強するために使用され得る。Gd含有造影剤は、腎性全身性線維症もしくは腎性線維化性皮膚症として公知の重篤な合併症が生じ得るので、腎機能が損傷した患者において使用できなかった。このような患者の必要性に対処するために、代替の画像化剤が必要とされる。金属をキレート化する化合物はまた、分子レベルでの詳細を検討するために、PET画像化およびSPECT画像化において使用される。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】米国特許第7,208,138号明細書
【特許文献2】米国特許出願公開第2004/0018974号明細書
【発明の概要】
【課題を解決するための手段】
【0007】
(要旨)
本開示は、一般に、トリペプチドモチーフおよびこのようなモチーフを使用するための方法に関する。これらペプチドは、金属に結合する能力を有し、このことは、これらペプチドを種々の適用に有用にする。特に、本開示のトリペプチドは、画像化、研究、化学療法、およびキレーション療法(chelation therapy)における適用を有する。
本発明は、例えば以下の項目を提供する。
(項目1)
配列XC1C2を有するトリペプチドを含む組成物であって、ここでXは、XC1C2が平面四角形配向もしくは四角錐配向またはその両方において金属を結合し得るような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、組成物。
(項目2)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目1に記載の組成物。
(項目3)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目1に記載の組成物。
(項目4)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目1に記載の組成物。
(項目5)
配列XC1C2を有するトリペプチドおよび金属を含む組成物であって、ここで該金属は、該トリペプチドと錯体化しており;そしてここでXは、トリペプチドおよび金属が平面四角形配向もしくは四角錐配向またはその両方を有する錯体を形成するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、組成物。
(項目6)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目3に記載の組成物。
(項目7)
前記金属は、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeからなる群より選択される、項目3に記載の組成物。
(項目8)
前記金属は、放射性同位体である、項目3に記載の組成物。
(項目9)
前記金属は、62Cuである、項目3に記載の組成物。
(項目10)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目3に記載の組成物。
(項目11)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目3に記載の組成物。
(項目12)
金属と、配列XC1C2を有するトリペプチドとを錯体化して、金属−XC1C2錯体を形成する工程を包含する方法であって;ここでXは、金属−XC1C2錯体が、平面四角形配向もしくは四角錐配向またはその両方を有するような任意のアミノ酸であり;そして
ここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、方法。
(項目13)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目12に記載の組成物。
(項目14)
前記金属は、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeからなる群より選択される、項目12に記載の組成物。
(項目15)
前記金属は、放射性同位体である、項目12に記載の組成物。
(項目16)
前記金属は、62Cuである、項目12に記載の組成物。
(項目17)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目12に記載の組成物。
(項目18)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目12に記載の組成物。
(項目19)
前記金属は、生物学的サンプル中に存在する、項目12に記載の方法。
(項目20)
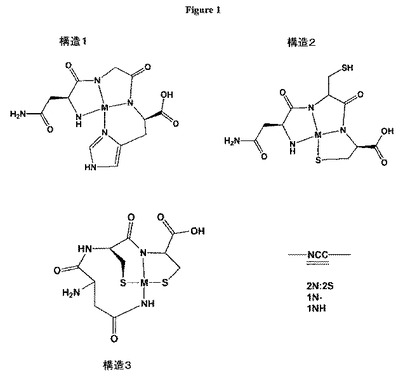
前記金属は、被験体において見いだされる、項目12に記載の方法。
(項目21)
前記金属−XC1C2錯体を検出する工程をさらに包含する、項目12に記載の方法。
(項目22)
前記金属−XC1C2錯体を画像化する工程をさらに包含する、項目12に記載の方法。
【0008】
特定の実施形態によれば、本開示は、配列XC1C2を有するトリペプチドを含む組成物を提供し;ここでXは、XC1C2が平面四角形配向もしくは四角錐配向またはその両方において金属を結合し得るような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0009】
他の実施形態によれば、本開示は、配列XC1C2を有するトリペプチドおよび金属を含む組成物を提供し;ここで上記金属は、上記トリペプチドと錯体化し;そしてここでXは、トリペプチドおよび金属が、平面四角形配向もしくは四角錐配向またはその両方を有する錯体を形成するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0010】
他の実施形態によれば、本開示は、金属と、配列XC1C2を有するトリペプチドとを錯体化して、金属−XC1C2錯体を形成する工程を包含する方法を提供し;ここでXは、金属−XC1C2錯体が、平面四角形配向もしくは四角錐配向またはその両方を有するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0011】
本発明の特徴および利点は、以下の実施形態の説明を読めば、当業者に容易に明らかである。
【0012】
(図面)
本開示のより完全な理解は、添付の図面とともに、以下の詳細な説明を参照することによって得られ得る。
【図面の簡単な説明】
【0013】
【図1】図1は、構造1 4N配位を有するGGH様金属錯体NGHを示す。構造2および構造3 いくつかの脱プロトン化骨格N原子によって上記金属の配位に影響を及ぼすGGH様配置を示す。
【図2】図2は、ヒトPRL酵素(PRl−1、PRL−2、およびPRL−3を含む)のC末端部分の配列アラインメントを示す。PRL−1のH166は、青で示されるNGHモチーフの一部である。PRL−1の残基C170およびC171は、赤で示されるCaaXモチーフ(ボックスで囲った)の一部である。上記NCCモチーフは、太字が付されている。
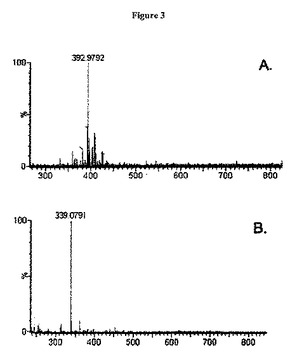
【図3】図3は、A. pH10において獲得したニッケル結合トリペプチドのESI−MS(ここで質量は、ニッケルが上記ペプチドに結合されていることを示す)、B.上記サンプルのpHを5に低下させた後のESI−MSスペクトル(ニッケルが、上記ペプチドから放出されたことを示す)を示す。
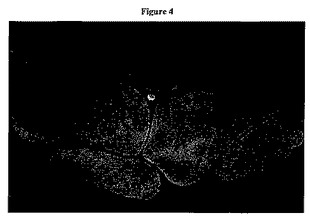
【図4】図4は、Cu−NCC磁気共鳴画像(MRI)を示す。
【図5】図5は、His−タグの切断および除去後に、Ni(II)(pH7.4)の存在下で精製した、精製WT PRL−1の写真を示す。写真は、その特徴的なさび色を示す。
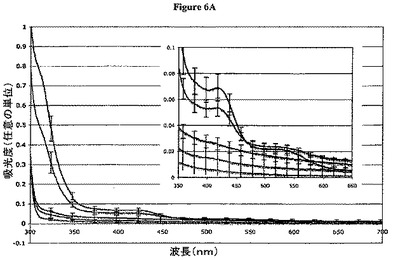
【図6A】図6Aは、Ni2+イオンの存在下で精製したPRL−1改変体の電子吸収スペクトルを示す。各タンパク質改変体について3つの(2つの)スペクトルを800〜200nmまで記録し、平均した。全ての改変体についてのタンパク質濃縮物は、A280比較に基づいて、5mg/mLから10%以内までである。700〜300nmまでのスペクトルを示す。目に見える特徴は、318nmおよび421nmでの吸収極大である。挿入図。421nmおよび526nmにおいて吸収極大の可視化のために、より小さなスケールで、600〜400nmまでのスペクトルが示される。曲線は、以下のとおりである:PRL−1−WT、黒(一番上);PRL−1−H166A、赤(一番上から2番目);PRL−1−C170S−C171S、橙色(一番下);PRL−1−C170S、緑(一番下から2番目);PRL−1−C171S、青(真ん中)。
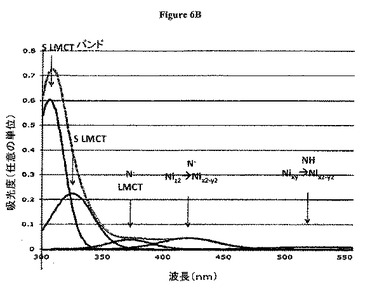
【図6B】図6Bは、PRL−1 WTの分解スペクトル(Resolved spectrum)。300〜700nmの間のNi精製PRL−1 WTタンパク質の上記UV−Visスペクトルの分解は、5つの曲線を使用するデータと適合する。その318nmのショルダーは、306nm、325nm、および372nmにおけるピークに分解される。上記分解された吸収ピーク(灰色破線)の和および生データ(黒実線)が、ともに示される。421nmおよび526nmにおけるピークもまた、存在する。
【0014】
特許および特許出願ファイルは、少なくとも1部のカラーで仕上げられた図面を含む。カラー図面付きのこの特許もしくは特許出願公開のコピーは、要求および必要な料金を支払えば、特許庁によって提供される。
【0015】
本開示は、種々の改変および代替の形態に影響されやすいが、特定の例示的実施形態は、図面に既に示されており、本明細書により詳細に記載される。しかし、特定の例示的実施形態の説明が、本発明を開示される特定の形態に限定すると意図されないが、一方で、この開示は、添付の特許請求の範囲によって規定されるような全ての改変および等価物を網羅することが理解されるべきである。
【発明を実施するための形態】
【0016】
(説明)
本発明は、一般に、短いペプチドモチーフおよびこのようなモチーフを使用するための方法に関する。これらペプチドは、選択的に金属に結合する能力を有し、このことは、これらペプチドを、種々の適用に有用にする。特に、本発明のペプチドは、画像化、研究、化学療法、およびキレーション療法における適用を有する。
【0017】
一般に、本開示は、選択された金属と強く結合する短い新規なペプチドモチーフ(MAPタグといわれる)に関する。よって、これらMAPタグは、とりわけ、上記選択された金属を、組成物から抽出するために使用され得る。本開示のMAPタグは、3アミノ酸長であり、N末端、C末端、もしくはその間の任意の位置において、より長いポリペプチドおよびタンパク質の中に含まれ得る。特定の実施形態において、しかし、上記MAPタグが、金属と結合するための上記MAPタグを表す、ポリペプチドもしくはタンパク質の配置に存在する(例えば、外部ループに存在する)ことは、有利であり得る。上記MAPタグはまた、非ペプチド実体に結合され得る。さらに、1種より多いMAPタグは、特定の分子上に存在し得る。
【0018】
多くのタンパク質が金属に結合することは公知である。しかし、このような結合は、代表的には、密な結合を達成し、次いで、開いた際に、容易に上記金属を放出するために、折りたたまれたタンパク質構造内で不連続なアミノ酸配列を利用する。他方で、MAPタグによる金属結合は、非常に密接して近くにある原子を使用して達成され、よって、極端な条件が、上記金属を放出するために必要とされる。上記タンパク質の熱変性および化学的変性は、上記金属のゆっくりとした放出を可能にする。例えば、極端な条件の使用(例えば、沸点温度、強酸)は、一定期間にわたって(例えば、数時間から長時間)上記金属をゆっくりと放出することをもたらし得る。
【0019】
(A.化学的構造)
特定の実施形態において、本開示は、新規なペプチドベースのタグもしくは金属の結合剤(MAPタグ)を提供する。上記MAPタグは、ポリペプチドもしくはタンパク質中に高親和性金属結合部位を直接コードするために使用され得る。本開示のMAPタグは、平面四角形および/もしくは四角錐の幾何的配置において金属に結合し得るそれらの能力によって特徴付けられる。
【0020】
(1.ペプチド配列)
本開示のMAPタグは、一般に、金属に結合し得る、少なくとも3個連続するアミノ酸残基を含む。本開示のMAPタグは、一般に、XC1C2によって表される配列を有し、ここでC1およびC2は、同じであってもよいし、異なっていてもよく、システイン、もしくはシステイン様非天然アミノ酸(例えば、硫黄含有α−アミノ酸もしくはβ−アミノ酸)であり得、そしてここでXは、上記形成されるペプチドタグが、平面四角形/四角錐の幾何的配置において金属に結合し得る限りにおいて、別の天然アミノ酸もしくは非天然アミノ酸、またはアミノ酸アナログであり得る。本明細書において使用される場合、L鏡像異性アミノ酸の略語は、従来どおりであり、以下のとおりである:
【0021】
【表1】

いくつかの実施形態において、Xは、アスパラギン(N)、グルタミン(Q)。ヒスチジン(H)、リジン(K)、もしくはアルギニン(R)であり得る。
【0022】
アミノ酸配列が、一連の三文字アミノ酸略語もしくは一文字アミノ酸略語として表される場合、標準的な使用および慣例に従って、左側がアミノ末端方向であり、右側がカルボキシ末端方向であることが理解される。
【0023】
本開示のMAPタグは、任意の組換え技術システムを使用して、発現のための遺伝子もしくはヌクレオチド配列と一続きで(in line)コードされ得る。さらに、上記タグは、ペプチドもしくはタンパク質の生成のための任意の合成法もしくは生合成法を使用して、ペプチドもしくはタンパク質へと組み込まれ得る。適用において、上記MAPタグは、金属と自発的に反応して、ペプチド−金属錯体を形成する。このようなペプチド−金属錯体は、溶液中で、または金属交換反応(transmetallation)もしくは任意の他のプロセスを介して、形成し得る。
【0024】
上記MAPタグは、単独で使用され得るか、あるいはN末端側、C末端側に結合され得るか、またはポリペプチド鎖内の可撓性リンカーもしくはループへと挿入され得るタグとして、使用され得る。上記MAPタグは、金属配位に関与する側鎖以外の任意の位置で改変され得る。よって、上記MAPタグは、Z−XC1C2−Z1のような配列を含み得、Zは、任意のアミノ酸もしくは任意のアミノ酸配列であり得、Z1は、Zに等しいもしくは等しくない、任意のアミノ酸またはアミノ酸配列であり得る。上記アミノ酸は、上記表1と同様に示される。非天然のおよびアミノ酸のアナログは、上記表中には列挙されていないが、それらは、Z、Z1、および/もしくはXに組み込まれ得る。
【0025】
特定の実施形態において、上記MAPタグは、別の分子に結合され得る。例えば、上記MAPタグは、炭水化物(例えば、ヒアルロン酸)のような非ペプチド実体に結合され得る。上記結合は、共有結合であってもよいし、リンカーを介して行われていてもよい。
【0026】
いくつかの実施形態において、上記MAPタグは、以下のような配列を含み得る:NC1C2;Z−NC1C2−Z1;Z−NC1C2;NC1C2−Z1;QC1C2;Z−QC1C2−Z1;Z−QC1C2;QC1C2−Z1;HC1C2;Z−HC1C2−Z1;Z−HC1C2;HC1C2−Z1;KC1C2;Z−KC1C2−Z1;Z−KC1C2;KC1C2−Z1;RC1C2;Z−RC1C2−Z1;Z−RC1C2;もしくはRC1C2−Z1。上記のように、Zは、任意のアミノ酸もしくは任意のアミノ酸配列であり得、Z1は、またはZに等しいかもしくは等しくない、任意のアミノ酸もしくはアミノ酸配列であり得る。上記アミノ酸は、表1と同じように表される。非天然のおよびアミノ酸のアナログは、上記表中には列挙されていないが、それらは、化合物ZおよびZ1に組み込まれ得る。
【0027】
一実施形態において、上記MAPタグは、以下の構造式を有するNCCを含む:
【0028】
【化1】
特定の実施形態において、本開示のMAPタグは、金属とのMAPタグ錯体化前、もしくは金属との錯体化後のいずれかに、上記MAPタグの標的化送達を提供する遺伝子もしくはヌクレオチド配列と一続きでコードされ得る。このことは、遺伝子、ペプチド、もしくは標的化に有用であることが公知の他のモチーフを使用して、達成され得る。例えば、MAPタグは、抗体、増殖因子、ペプチドなどと一緒に組み込まれ得る。
【0029】
(2.金属結合)
特定の実施形態において、本開示のMAPタグは、単独で、またはポリペプチドもしくはタンパク質に組み込まれる場合、金属と錯体化して、平面四角形/四角錐の幾何的配置を有するMAPタグ−金属錯体を形成し得る。上記金属は、上記MAPタグと、2N:2S配位を介して錯体化し得る。上記MAPタグはまた、当業者によって認識される適切な条件下で、平面四角形の幾何的配置において多くの金属に結合し得る。一般に、本開示のMAPタグは、IUPAC Groupを参照すると:3族金属(例えば、Y);5族金属(例えば、VおよびU);6族金属(例えば、Cr、Mo、W);7族金属(例えば、Mn、Tc、Re);8族金属(例えば、FeおよびRu);9族金属(例えば、Co、Rh、Ir);10族金属(例えば、Ni、Pd、Pt);11族金属(例えば、Cu、Ag、Au);12族金属(例えば、Zn、Cd、Hg);13族金属(例えば、Al、Ga、In、Tl);14族金属(例えば、SnおよびPb);ならびに15族金属(例えば、Bi)を結合し得る。特定の実施形態において、上記MAPタグは、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeと結合しかつMAPタグ−金属錯体を形成し得る。
【0030】
上記のように、上記金属を結合すると、上記MAPタグは、平面四角形の幾何的配置を有する。本開示はまた、種々の平面四角形配置にある、MAPタグ金属配位を企図する。例えば、上記MAPタグがNCCでありかつ上記金属が上記のようにMとして表される任意の金属である場合、上記MAPタグは、式1によって表される構造式を有し得る:
【0031】
【化2】
他の構造的配置がまた、企図される。例えば、上記MAPタグがNC1C2でありかつC1が非天然アミノ酸(例えば、(S)−2−アミノ−2−メルカプト酢酸)でありかつC2がシステインである場合、上記MAPタグの構造式は、式2として表され得、ここでMは、上記のように任意の金属である:
【0032】
【化3】
別の例において、上記MAPタグは、NC1C2であり、C2は、非天然アミノ酸(例えば、(S)−2−アミノ−4−メルカプト酪酸であり、上記MAPタグの構造式は、式3として表され得、ここでMは、上記のように任意の金属である:
【0033】
【化4】
別の例において、上記MAPタグは、NC1C2であり、C2は、βアミノ酸(例えば、3−アミノ−2−メルカプトプロピオン酸であり、上記MAPタグの構造式は、式4として表され得、ここでMは、上記のように任意の金属である:
【0034】
【化5】
別の例において、上記MAPタグは、RCCであり、上記MAPタグの構造式は、式5として表され得、ここでMは、上記のように任意の金属である:
【0035】
【化6】
別の例において、上記MAPタグは、QCCであり、上記MAPタグの構造式は、式6として表され得、ここでMは、上記のように任意の金属である:
【0036】
【化7】
(B.上記MAPタグの適用)
(1.金属抽出のためのMAPタグ)
上記のように、MAPタグは、高親和性で金属を結合し得る。MAPタグはまた、液体から固体までの範囲に及ぶ種々の組成物から金属を抽出し得る。結論として、MAPタグが、配位を共有するのではなく上記金属を抽出する能力は、以下で依り詳細に記載されるように、上記タグを、特定の金属を別の組成物から分離することにおける使用に影響を受けやすくする。上記MAPタグは、上記金属と錯体化し、次いで、上記金属を組成物中の成分(例えば、キレート化剤(例えば、EDTA)、または例えば、IDAもしくはNTAと結合体化した固体支持体)から上記金属を抽出もしくは除去することによって、上記組成物から金属イオンを封鎖する。よって、上記MAPタグは、金属抽出ペプチド(MAP)タグである。
【0037】
さらに、少なくとも一部は、MAPタグが金属に選択的に結合する能力に起因して、本開示の実施形態は、キレーション療法に有用であり得る。従って、MAPタグは、生物学的に毒性の金属(例えば、鉛)を封鎖するために使用され得る。
【0038】
同様に、MAPタグはまた、金属を水、他の水溶液、もしくは身体の外の任意の組成物から封鎖することにおいて有用であり得る。上記MAPタグ自体は、一般に、多くの溶媒中で可溶性である。しかし、特定の実施形態において、上記MAPタグ−金属錯体は、多くの有機溶媒中では溶解性が乏しく、いったん金属結合が起こると、沈殿し得る。このことは、有機溶媒からの抽出の間の効率的な分離を可能にする。
【0039】
(2.画像化適用)
特定の実施形態において、本開示は、MAPタグを使用する、画像化のための方法を提供する。このような方法は、治療的画像化および医療的画像化、または他の画像化適用のために使用され得る。上記MAPタグと錯体化するように選択される金属は、上記特定の適用および画像化技術に依存する。例えば、常磁性金属は、画像化技術(例えば、MRI、NMRおよびEPR)との使用に選択され得る一方で、金は、電子顕微鏡のような技術のために選択され得る。その一方で亜鉛は、蛍光のような適用のために選択され得る。どの金属が所定の適用および画像化技術に適しているかは、当業者によく知られている。
【0040】
一実施形態において、MAPタグは、画像化技術において使用するための試薬を調製するために利用される。すなわち、MAPタグは、金属に結合し、続いて、画像化のために使用され得る(例えば、上記金属が画像化のために使用され得るように、被験体の身体において使用され得る)錯体を提供する。適切な画像化技術の例としては、単独でもしくは組み合わせにおいて、コンピューター断層撮影法(CT)、磁気共鳴画像法(MRI)、超音波、陽電子放射断層撮影法(PET)、単一光子放出型コンピューター断層撮影法(SPECT)、核磁気共鳴法(NMR)、電子常磁性共鳴(EPR)、電子顕微鏡、蛍光画像法などが挙げられる。
【0041】
別の実施形態において、MAPタグは、被験体の一部に局在するポリペプチドもしくはタンパク質、または非タンパク質実体へと組み込まれ得る。ここでMAPタグ−金属錯体の金属部分は、局在部位(例えば、組織もしくは腫瘍)を画像化する能力を提供する。例えば、上記MAPタグ−金属錯体は、上記MAPタグ化分子の標的化部分に対するレセプターおよび/もしくは結合パートナーを発現する組織もしくは他の構造を同定するために使用され得る。上記金属を可視化し得る画像化技術は、このような特徴の同定のために使用され得る。
【0042】
例えば、MRI造影剤は、隣り合う原子の緩和を変化させて、上記造影剤が存在する領域と、存在しない領域との間で検出されるシグナルの差異を発生させる、代表的には、金属中心を含む常磁性種である。MAPタグは、Cuと錯体化される場合には、常磁性であり得る。従って、MAPタグ−金属錯体は、MRI造影剤として使用するための能力を有する。大部分の造影剤は、顕微鏡レベルで標的化されないが、身体もしくは組織中の肉眼的構造および欠損を明らかにするために使用される。適用のこのタイプにおいて、上記造影剤の濃度は、非常に高い。造影剤を、選択された組織もしくは顕微鏡的構造に標的化することは、細胞レベルでの異常の同定を可能にする点で有益である。例えば、上記MAPタグは、特定のレセプターを発現する癌細胞を認識する抗体上のタグとして発現され得る。標的化された送達は、より低い濃度の造影剤が投与されることを可能にする。このことは、とりわけ、毒性および他の副作用を最小にする助けになり得る。
【0043】
62Cuが、身体内の分子的な詳細を画像化し得る、PETおよびSPECT画像化において使用される放射活性同位体であることは公知である。よって、MAPタグは、Cu(もしくは別の金属)の適切な放射性同位体を添加され得、これら画像化技術でのトレーサーとして使用され得る。同様に、上記タグを、任意のタンパク質もしくはペプチド、または細胞の部分セットを結合する他の分子に融合することによって、標的化された画像化が、達成され得る。
【0044】
上記MAPタグは、特定の金属に対して高い親和性を有する短い配列であるので、とりわけ有利である。従って、発現されたタンパク質もしくは他の分子においてMAPタグをコードすることによって、生物内の特定の細胞タイプへの標的化された送達は、可能である。さらに、細胞の部分セットを標的化することは、画像化に必要とされる造影剤の量の減少を可能にする。
【0045】
(3.化学療法)
本発明はまた、化学療法を補助するために使用され得る。例えば、MAPタグは、白金および/もしくは放射性核種との錯体を形成し得、特定の細胞もしくは細胞タイプへの標的化された送達を提供するために使用され得る。
【0046】
本発明のよりよい理解を促進するために、以下の特定の実施形態の実施例が提供される。以下の実施例が、本発明の範囲全体を限定するようにも、規定するようにも、決して解釈されるべきではない。
【実施例】
【0047】
(実施例1)
変異誘発技術を、選択された金属と強い結合親和性を有する上記ペプチドモチーフを同定するために使用した。簡潔には、ヒトPRL−1遺伝子を、pET−30 Xa LICベクター(Novagen)にクローニングし、PCRベースのQuikChange法(Stratagene)を使用して変異させた。C170S、C171S、およびH166A変異体を生成するプライマー(5’から3’)は、それぞれ、
【0048】
【数1】
および
【0049】
【数2】
(Integrated DNA Technologies,Coralville,IA)であった。大文字は、変異した塩基を示す。PCR反応物をDpnI(Promega)で、1.5時間にわたって37℃において処理し、NovaBlue GigaSingles Competent細胞(Novagen)に直接形質転換し、30mg/mL カナマイシン選択でLB上にまいた。個々のコロニーを、選択的LB中で37℃において一晩増殖させた。得られたDNAを、Wizard Plus MiniPrep System(Promega)を使用して精製した。全ての変異を、DNA色素−ターミネーター配列決定によって確認した(Northwoods DNA,Inc.,Bemidji,MN)。上記二重変異体C170S/C171Sを、第2回の変異誘発によって生成し、同様に確認した。
【0050】
PRL−1および関連する変異遺伝子を有するベクターを、BL21(DE3)細胞(Novagen)へと形質転換し、選択的LB上で37℃において一晩増殖させた。選択されたコロニーを、選択的M9ZBブロス中で、一晩37℃において増殖させた。次いで、一晩の培養物を、500mLの非標識最小培地に移し、0.6〜0.8の間のOD550において、1mM IPTGで誘導した。NMR実験のために発現させたタンパク質は、15N−塩化アンモニウムを含む最小培地中で増殖させた。上記組換えタンパク質の過剰発現を、SDS−PAGEによって確認した。細胞を、4400×gでの遠心分離によってペレット化し、使用するまで−80℃で保存した。
【0051】
一連のPRL−1変異体を、金属配位に関与する残基を同定するために試験した。GGH様モチーフを含むモデルペプチドは、極めて高い親和性でNi(II)およびCu(II)を配位することを示した。このモチーフにおいて、2つのGly位置のうちのいずれかにおける置換が、ごくわずかに結合を低下させるのに対して、Hisの変異は、結合を破壊する。PRL−1は、NGH、GGH様コンセンサスモチーフを、そのC末端付近においてコードする(図2)。従って、PRL−1−H166Aを、PRL−1に含まれる上記NGH配列が、金属結合を担うという仮説を試験するために作製した。PRL−1およびPRL−1−H166Aを、それぞれ、約120μMおよび60μMのNiを結合したICP−MSによって、並行して分析した。吸収分光法によって測定した場合に、わずか20%のシグナルの低下が、H166Aについて認められた。タンパク質シグナルが、等しい程度にまで減少され、可視領域のシグナルの損失が、タンパク質不安定性に起因することはまた、吸収スペクトルから明らかである。タンパク質不安定性は、上記2つの検出法における不均衡が原因のようである。なぜなら、ICP−MSは、分析する前に、実質的により多くのサンプル操作を要するからである。上記ヒスチジン変異体は、堅い結合を保持したままであったので、C170および/もしくはC171における変異を、チオールがNi配位にしばしば関与することから、作製した。個々のシステイン変異体、ならびに二重変異体におけるNiレベルは、野生型タンパク質と比較して、大いに低下した。このことは、両方のCys残基が、金属配位に影響を及ぼすことを示す。C170S変異体においても、上記C170S−C171S二重変異体においてもNiが検出されなかったが、約17μMの小さなシグナルが、PRL−1−C171Sについて認められた。
【0052】
(実施例2)
タンパク質精製技術を使用して、上記タンパク質−金属錯体を精製した。簡潔には、細胞ペレットを、30mL 緩衝液A(100mM NaCl、50mM Tris−HCl、10mM イミダゾール、pH7.4、Ar散布した)中に再懸濁し、15,000psiにおいて、フレンチプレッシャーセル(ThermoElectron)を使用して溶解した。緩衝液のアルゴン散布を行って、酸素を置換した。次いで、サンプルを、21,000×gにおいて遠心分離し、その上清を、0.2μm ナイロンフィルタを通して濾過した。(His)6−タグ化PRL−1の精製を、流速1mL/分において、Akta Explorer精製システムを使用して行った。サンプルを、金属添加した5ml HiTrap Chelating Column(GE Healthcare)に載せ、載せた後に、5×カラム容積ですすいだ。溶出を、13×カラム容積かけて60% 緩衝液B(100mM NaCl、50mM Tris−HCl、500mM イミダゾール、pH7.4、アルゴン散布ありもしくはなし)へ、3×カラム容積かけて100% Bへの直線勾配を使用して達成し、および4×カラム容積かけて100% Bにおいて維持した。溶出を、280nmでの吸収によってモニターし、画分を、SDS−PAGEを使用して純度について試験した。適切な画分をプールし、Ar散布したもしくはAr散布していない100mM NaCl、50mM Tris−HCl、pH7.4に対して透析した。上記(His)6−タグを、第Xa因子プロテアーゼ(Promega)を、室温において一晩使用して、上記標的タンパク質から切断した。XarrestTMアガロース(Novagen)を使用して、プロテアーゼを除去し、上記カラムに再度載せて上記タグおよび任意の非切断タンパク質を除去する前に0.2μm ナイロンフィルタで濾過した。上記タンパク質を、透析緩衝液中の上記カラムに載せ、最初のフロースルーを集めて、上記タンパク質サンプルを作製した。サンプルを濃縮し、(Amicon Ultra)10kDa MWCO遠心分離フィルタを使用して、Ar散布100mM NaCl、50mM TrisCl、pH7.4中で、少なくとも106倍交換した。上記サンプルの純度および変異体確認を、それぞれ、SDS−PAGEおよびESI−MSによって確認した。カラムを、1M NaOHで処理し、2×カラム容積の100mM EDTAでストリッピングし、Ar散布した100mM NiSO4、100mM CuSO4、100mM CuCl2もしくは100mM ZnCl2を、各使用の直前に再度充填した。
【0053】
図5は、上記His−タグの切断および除去後の精製PRL−1を示す。上記his−タグに結合した金属は、八面体の幾何的配置を有し、これは、青緑色であり、このような配位事象に対応する吸収は、スペクトルのうちの700〜800nm領域において現れる。Ni(II),pH7.4の存在下で精製したWT PRL−1の写真は、その特徴的なさび色を示す。上記精製した野生型PRL−1タンパク質は、予測外のさび色を有する(図5)。このことは、芳香族残基(すなわち、TrpもしくはTyr)の金属錯体化もしくは酸化から生じ得る。
【0054】
芳香族残基の酸化は、上記タンパク質のインタクトなおよびトリプシン消化研究からの質量分析データに基づいて容易に排除された。さらに、本発明者らは、上記さび色のタンパク質が、高速遠心分離の間にチューブの底で濃縮されるので、上記タンパク質の密度が、予測されるより高いことを観察した。金属配位が上記タンパク質錯体の密度を増加させるので、本発明者らは、上記発色団が、上記タンパク質との金属錯体の形成によって生成される可能性をさらに調べた。金属キレート化クロマトグラフィーを使用して、PRL−1を精製したので、金属イオンのもっとも可能性のある供給源は、上記IMAC樹脂に由来するNiであった。
【0055】
組換えWT PRL−1を含む細胞溶解物が、等しく分けて、Ni2+充填した、Cu2+充填したもしくはZn2+充填したIMAC樹脂を使用して並行して(side−by−side)精製した場合、上記タンパク質は、上記キレート化樹脂に結合した金属を抽出する。金属交換反応の速度は、ニッケルで最も速く、続いて、銅であった。亜鉛は、全てのサンプルにおいてある程度存在し、このことは、上記タンパク質が亜鉛をインビボで拾い上げることを示す。亜鉛が上記カラムに結合される場合、亜鉛のみが、上記タンパク質中で認められる。Apo PRL−1は、可溶性が乏しく、金属の損失が沈殿をもたらす。誘導結合型プラズマ質量分析法(ICP−MS)によって試験した場合に各精製からの可溶性物質は、結合した金属の等しい割合を有する。
【0056】
(実施例3)
ICP−MSを使用して、上記結合した金属を特徴付けした。簡潔には、精製タンパク質サンプルを、PFA微小遠心分離管(Savillex,#s 7240、7241)において濃硝酸中、68±1℃で16.5±0.7時間にわたって消化した。消化前のタンパク質定量に基づいて、次いで、上記サンプルを、20mM 炭酸水素アンモニウム(pH8.2)中で1.5μMへと希釈し、0.2μm ナイロンフィルタを通して濾過し、微小同軸ネブライザ(microconcentric nebulizer)を装備したVG Elemental VGII+XS Inductively−Coupled Plasma−Mass Spectrometerへと注入した。最小で、2つの別個に調製したサンプルを、各タンパク質改変体について分析した。各サンプルを2回スキャンし、標準物質を、各サンプルスキャンの前、その間、およびその後に走らせた。ドリフトを、サンプルの注入と標準物質の注入との両方のスキャンの間の洗浄している間にモニターした。
【0057】
上記発色団が、荷電したIMACマトリクスからの金属のタンパク質抽出から生じるという本発明者らの仮説を調査するために、サンプルを、半定量的分析のためにICP−MSに供した。バックグラウンド金属を除去するために激しい洗浄を使用したので、この方法から得られた値は、結合した金属量より本質的に小さく見積もられるが、上記タンパク質に対して異なる金属の相対的親和性を評価するために使用され得る。さらに、上記細胞における発現の間にPRL−1に結合してしまった可能性のある内因性の金属リガンドを同定するために、ICP−MSを使用して、周期表中の最初の2列の遷移金属、および他の一般的に生物学的に関連する金属の存在を探した。顕著なことには、上記データは、Znおよび特定の金属(例えば、Ni)にのみ結合したPRL−1が、精製の間に上記IMACカラムで使用されたことを示した。16時間にわたって70℃において硝酸中で消化した、平均して500μM Ni精製タンパク質サンプルは、約180μM Niを含むと決定され、これは、1:2.7+/−0.7のNi 対 タンパク質化学量論を生じた。Znはまた、約60μMにおいて上記Ni精製サンプル中で検出された。別個に、Zn−IMACを使用するPRL−1の精製は、Znが、270μMにおいて多い量で存在する唯一の金属であり[1 Zn:1.8 タンパク質]、これらタンパク質サンプルが、Niのわずかなバックグラウンドレベルを含むことを示した。Ni精製PRL−1およびZn精製PRL−1の両方のICP−MSは、顕著な量で他の金属を示さず、このことは、亜鉛が、インビボで配位され、上記ニッケルのクロマトグラフィー精製の間に、Ni2+によって部分的に置換されたことを示唆する。ICP−MS分析は、Ar流中で行われ、Arの最も量の多い同位体がCaと同じ質量を有するので、Ca結合は、一次質量シグナル(primary mass signal)からは決定できない。にも関わらず、遙かに多い量のCaが、Znと比較すると細胞培養培地中に存在し、それでもなお上記ICP−MSシグナルが、主に亜鉛に対応するので、上記タンパク質は、カルシウムに結合しないようである。上記ICP−MSデータは、PRL−1がNi−IMACを使用して精製される場合、Znが評価できる程度の量で存在することを示す。このことは、Zn結合が細胞において存在することを示唆する。
【0058】
(実施例4)
UV−Vis吸収スペクトルを、上記PRL−1タンパク質の濃度を測定するために使用した。簡潔には、精製PRL−1アナログの濃度を、280nmにおいて測定し、19420M−1cm−1の吸光係数を使用して計算した。高い200nm領域での考えられる金属−タンパク質吸収干渉が原因で、上記UV−Vis定量法の有効性を、Bradford法によって確認した。10mg/mL、5mg/mL、2.5mg/mL、および1.25mg/mLの濃度のPRL−1アナログのスペクトルを、800〜200nmまで、Cary 100 UV−Vis分光光度計で集めた。上記分析における散乱効果を較正するために、PRL−1−WTスペクトルを、上記PRL−1−C170S−C171Sスペクトルを差し引くことによって調整した。なぜなら、上記PRL−1−C170S−C171Sスペクトルは、金属を結合しないからである。得られたスペクトルを分解し、ピーク位置を、GRAMS/AI 7.00ソフトウェアを使用して評価した。3つの方法は、金属置換を目的とした。第1において、PRL−1−WT(例えば、野生型)サンプルを、20mM β−メルカプトエタノール(BME)を使用して還元し、イミダゾールを、10mMの濃度になるように添加した。0.5mM タンパク質サンプルに対して、これは、BMEの400倍溶液過剰に対応して、上記2つのC末端システインチオールと競合し、イミダゾールの200倍溶液過剰に対応して、単一のC末端ヒスチジンイミダゾール側鎖と競合する。新たなベースラインをとり、スペクトルを再計算した。第2の方法において、ベースラインを、100μM EDTAを含む緩衝液について再計算し、100μM EDTA中で1000万倍交換した上記PRL−1−WTサンプルに対するスペクトルを、記録した。最後に、10mg/mL タンパク質サンプルを、5.6M グアニジウム塩酸塩中で変性させ、90℃へと加熱し、新たなベースラインをとり、そのスペクトルを、0時間、2時間、4時間、および6時間で集めた。
【0059】
よって、PRL−1による金属配位の特徴を、UV−Vis吸収分光法で決定した。PRL−1およびその変異体が使用した上記金属配位幾何的配置を、800〜200nmにおいてUV−Vis吸収分光法を使用して分析した。PRL−1の吸収スペクトルは、280nmおよび220nmにおいてピークを生成した。このことは、上記タンパク質内の芳香族およびペプチド結合から生じ、上記Bradfordアッセイを使用して決定される濃度とよく一致する。金属含有化合物は、可視範囲においていくつかの波長でしばしば吸収し、上記Ni精製およびCu精製のPRL−1タンパク質はまた、可視範囲においてピークを表し、これは、上記サンプルのさび色に対応する。PRL−1 WTは、318nmにおいて最大(非常に大きな280バンドのショルダーとして明らか)、421nmにおいてより広いシグナル、および526nmにおいて非常に広いピークを示す(図6A)。Cu精製PRL−1は、Ni精製PRl−1と同様のスペクトルプロフィールを生じる(データは示さず);しかし、小さい青色のシフトは、上記個々のスペクトルピークに対して観察され、このことは、CuをNiの代わりに使用した場合に予測される。予測されるように、Znを充填した樹脂を使用して上記タンパク質を精製した場合には、吸収も目に見える色も認められなかった。このことは、Zn(II)が完結したd殻を有し、よって、可視スペクトルにおいて電子移動を示すはずがないという事実に起因する。
【0060】
上記WTタンパク質と比較して、上記H166A改変体は、同じ吸収プロフィールを示したが、上記ピークは、上記ICP−MS結果と一致して、ある程度強度が低下した。PRL−1−WTおよびPRL−1−H166Aのスペクトルは、可視領域において上記システイン変異体のものとは異なる。等しい濃度において、システイン170および/もしくはシステイン171におけるセリンでの個々の置換は、421nmおよび526nmでのシグナルの完全な損失を示す。これら変異タンパク質を、10mg/mL(0.5mM)と同程度の濃度において試験したところ、特異的吸収バンドは検出されなかった。上記WTからのシグナルは、0.1mg/mLにおいて明らかであり、このことは、Niの結合が、各変異によって、少なくとも100倍低下することを示す。
【0061】
較正したWTタンパク質の上記UV−Visスペクトルのデコンボルーションを、300nm〜700nmの間で行った。すると、318nmピークが、それぞれ、306nm、325nm、および372nmにおいて、全ての複合ピーク吸収に対して54%、38%および7%寄与する3つの別個の吸収バンドから構成されることが明らかになる(図6B)。上記PRL−1−H166Aの吸収プロフィールは、上記野生型タンパク質スペクトルと非常に似ている。残基166における上記イミダゾール基の非存在は、吸収極大をシフトもせず、318nm吸収の成分バンドに顕著に寄与もしない。このことは、上記NGHモチーフ内のヒスチジンに由来するイミダゾールが、Niを直接配位しないことをさらに示す。
【0062】
PRL−1におけるNi結合は、PRL−1の活性部位におけるジスルフィド結合の還元によって影響を受けない。20mM BMEの添加は、上記スペクトル吸収プロフィールのUVもしくは可視領域を変化させず、このことは、金属結合が、上記タンパク質の還元および上記還元剤の存在によって大きく影響を受けないことを示す。上記還元型スペクトルを、上記酸化型から差し引くと、上記還元型WTサンプルと、上記酸化型WTサンプルとの間には、金属配位に有意差がないことが確認される。Hisからの小さな貢献がなされるか否かを調べるために、さらなるイミダゾールを、上記溶液に添加して、一時的な会合のシグナル強度を増大させた。10mM イミダゾールの添加は、上記吸収プロフィールに対して影響がなかった。このことは、上記Niへの接近が制限されていることを示唆する。さらに、上記Ni精製サンプルが、イミダゾールの存在下で還元された場合に、スペクトル変化は認められなかった。Ni精製WTサンプルの、100μM EDTAを含む緩衝液への1000万倍の交換は、上記スペクトルに対してほとんど影響がなく、このサンプルに対する吸光度値は、上記WT吸光度の誤差範囲内であった。7.4から8.5へとpHを上げると、モル吸収係数(ε)を約6倍増大させるはずであるが、興味深いことには、pHが上昇した場合に、εの変化は認められなかった。このことは、上記結合したNiが、PRL−1に接近できない溶媒であることを示唆し得る。しかし、6.5へと上記pHを下げると、目に見えるピークの強度は低下する(データは示さず)。
【0063】
Ni結合は、pH7.4において不可逆性になるようである。なぜなら、Ni精製PRL−1に由来するNiの移動が、攻撃的な手段を要するからである。Ni精製PRL−1を、5.6M グアニジウム塩酸塩で90℃において処理した場合に、318nm、421nmおよび526nmにおける上記ピークは、6時間かけて徐々に消失するに過ぎない(データは示さず)。750nmにおけるシグナルの付随した出現は、6時間かけて観察された。このことは、Niが、上記タンパク質から放出され、八面体の幾何的配置において水によって配意されたことを示す。図6Aは、Ni2+イオンの存在下で精製したPRL−1改変体の電子吸収スペクトルを示す。各タンパク質改変体に対する3つのスペクトルを、800〜200nmで記録し、平均した。全ての改変体についてのタンパク質濃度は、A280比較に基づいて、5mg/mLから10%以内までである。700〜300nmのスペクトルが示される。目に見える特徴は、318nmおよび421nmにおける吸収極大である。図6Aにおいて、曲線は、以下のとおりである:PRL−1−WT、黒色;PRL−1−H166A、赤色;PRL−1−C170S−C171S、橙色;PRL−1−C170S、緑色;PRL−1−C171S、青色。
【0064】
図6Bは、PRL−1 WTの分解したスペクトルを示す。300〜700nmの間でのNi精製PRL−1 WTタンパク質の上記UV−Visスペクトルの分解は、5つの曲線を使用した上記データと適合する。上記318nmのショルダーは、306nm、325nm、および372nmにおけるピークに分解される。上記分解された吸収ピークの和(灰色破線)および生データ(黒色実線)が、両方示される。421nmおよび526nmにおけるピークも、存在する。
【0065】
図6Bの挿入図は、600〜400nmのスペクトルを示し、421nmおよび526nmにおける吸収極大の可視化のために拡大スケールで示される。
【0066】
(実施例5)
エレクトロスプレーイオン化質量分析法(ESI−MS)を使用して、上記サンプルを分析した。簡潔には、ESIスペクトルを、MSモードで操作したQ−Tof−2(Micromass Ltd,Manchester UK) Hybrid Mass Spectrometerで獲得し、感度について8000分解(FWHH)に離調した(de−tuned)フライトアナライザーの時間とともにデータを獲得した。全タンパク質ESIスペクトルについては、精製タンパク質サンプルを、未処理(「酸化」)20mM 炭酸水素アンモニウム(pH8.2)中で1μg/μLに希釈し、室温でインキュベートした。精製後1日目、4日目、および7日目の時点で採取し、使用する準備が出来るまで−80℃で凍結した。サンプルを、短い(3cm×1mm内径)逆相(RP)HPLCカラム(Hamilton PRP1,Reno,NV)で脱塩した。サンプルを、タンパク質(5μg)を含む1% ギ酸溶液から上記カラムに載せ、同じ溶液中で洗浄し、90% MeOH/0.5% ギ酸で、上記ESI源へと直接溶出した。コーン電圧(cone voltage)は、60eVであり、衝突セル上の電圧は、20Vであった。800〜3000uの質量範囲にわたってスペクトルを獲得し、1サイクルあたり5秒にわたってデータを蓄積した。
【0067】
トリプシン消化のために、PRL−1−WT、PRL−1−H166AおよびPRL−1−C170S−C171Sを、20mM β−メルカプトエタノールありまたはなしの炭酸水素アンモニウム中(pH8.2)で1mg/mLに希釈し、トリプシン(Promega)を、1:50比のプロテアーゼ:タンパク質(w/w)で添加し、37℃において16時間にわたってインキュベートした。完全な消化を確認するために、種々の時点でのサンプルをSDS−PAGEで泳動し、銀染色で可視化した。MS分析と組み合わせて、キャピラリーHPLC分離を、10μL/分での勾配を発生させるクロマトグラフ(Waters capLC XL,Milford,MA)とともに、Micro−Tech Scientific(Sunnyvale,CA)によって充填されたZorbax SBC18 RPカラム(5cm×0.32mm内径,3.5μm ビーズサイズ、300Å ポアサイズ)を使用して行った。20%から80% Bへの直線勾配を、120分かけて適用した。溶媒は、以下であった:A 99% H2O、1% MeOH、B 99% MeOH、1% H2O(ともに0.08% ギ酸)。アルゴンを、上記ビームを約20%まで減衰させる圧力で、上記衝突セルに入れた。このことは、供給調節器での16psiに、または衝突セル付近にあるペニングゲージでの5.3×10−5mBarに対応する。上記衝突セルを、最大移動のために8Vで操作し、スペクトルを、250〜2000uの範囲にわたって獲得し、1サイクルあたり8秒間のデータを蓄積した。
【0068】
NMR実験を行って、上記サンプルを分析した。簡潔には、2D 1H−15N HSQCスペクトルを、パルスフィールド勾配を備えた低温の三重共鳴(triple−resonance)プローブを使用して、Bruker Avance 800 MHz NMR分光器で獲得した。水抑制を、フリップ−バック(flip−back)パルスを使用して達成した。全てのスペクトルを、37℃において得、1Hにおいて2048ポイントでの16回のスキャンで獲得した。256増分もしくは128増分を、それぞれ、WTおよびC170S−C171Sの15Nにおいて集めた。サンプルを、50mM リン酸ナトリウム、100mM NaCl(pH6.5)中に調製し、5% D2Oを含めた。WT PRL−1およびPRL−1−C170S−C171Sの濃度は、それぞれ、1.0mMおよび0.8mMであった。サンプル還元を、スペクトル獲得の前に、10mM DTTを少なくとも24時間添加することによって達成した。1H化学シフトを、D2O中の外部DSS標準に関して参照した(29)。1Hに対する間接的な参照は、13Cおよび15Nに対して決定し、このことから、比13C/1H=0.251449530および15N/1H=0.101329118を想定した。コンピュータープログラムnmrPipeおよびSparkyを、データ処理およびスペクトル分析のために使用した。
【0069】
(実施例6)
上記MAPタグモチーフが金属結合を完全に担うことを確認するために、本発明者らは、NCCを合成した。上記NCCペプチドを合成し、精製し、上記ペプチドを、金属充填IMAC樹脂とともにインキュベートした。錯体形成を、陰イオンモードで作動するエレクトロスプレーイオン化質量分析法(ESI−MS)を使用して確認した(図3)。
【0070】
上記NCCペプチドにおいて、吸収シグナルは、PRL−1中に埋まった上記ペプチド−金属錯体について観察したことを模倣するが、上記錯体は、上記トリペプチドの中により露出している。上記可視範囲における吸収は、そのpHが6から7.4へと上昇する場合に強くなる。酸性化は、そのシグナル強度を減少させ、より塩基性のpHに戻った場合に、完全なシグナルが戻ることはない。このことは、上記平面四角形の幾何的配置からいくつかの金属の不可逆的な損失が起こり得ることを示す。上記pHを10へと上昇させると、第5の配位子である、おそらく水もしくはヒドロキシドが、配位し、四角錐配置を生じることが示唆される。pH7.4において、この配位子は、おそらく高度に交換可能であり、吸収分光法によって検出するにはあまりに一過性過ぎる。上記交換可能な配位子は、上記バルク溶媒の常磁性緩和への達成方法を提供し、MRIによって検出され得る(図4)。さらに、上記金属塩を、上記ペプチドを含む水溶液へと混合することによって、上記金属を添加した。条件に依存して、上記ペプチドの吸収スペクトルは、金属移動を介して形成された上記錯体に類似であったか、または全長タンパク質について得られたものとは全く異なっていたかのいずれかであった。上記異なるスペクトルは、上記金属が、この場合、硫黄によってのみ配位されている可能性があることを明らかにする。上記NCCペプチドは、金属結合を付与するに十分であるが、生成される錯体のタイプは、上記金属が上記ペプチドに導入される方法に依存する。金属交換反応は、上記金属を、上記NCCトリペプチドに組み込んで、極めて高親和性の平面四角形の錯体を生成するための効率的な方法を提供する。
【0071】
NGHのGGH様金属錯体は、図1の構造1に示される。本発明者らは、上記金属結合NCC錯体について2つの考えられる化学的構造を想定した。これらを、図1の構造2および構造3に示す。構造2は、いくつかの脱プロトン化骨格N原子によって上記金属の配位を伴うGGH様配置に対応する。この場合、Hisに由来するイミダゾリウム窒素は、NCCにおいて、配位した錯体における3位でCys由来の硫黄と交換される。この錯体は、3N:1S配位を有し、このことは、本発明者らの吸収スペクトルとは一致しない。上記NCCによる独特の特異的配位を確認するために、NGCも合成し、金属結合における中心的Cys側鎖の重要性を確認した。上記IMAC樹脂とともにインキュベートした上記NGCペプチドは、上記金属結合したNCCもしくはPRL−1スペクトルと似た吸収スペクトルを生じない。よって、本発明者らは、GGH様配位が利用されず、上記特定の幾何的配置において金属を結合するために遙かに低い親和性を有するはずであると結論づけた。
【0072】
構造3は、上記ペプチド単独、およびより大きなタンパク質の状況にある上記ペプチドの両方について得られたデータと完全に一致している。両方についての上記吸収スペクトルは、2つに硫黄原子が、2つの窒素原子とともに上記錯体に関与していることを示し、上記窒素原子のうちの一方は脱プロトン化され、他方は、1つのプロトンが結合されている。構造3は、N末端アミン基を伴わなくてもよく、このアミン基は、上記ペプチドおよびタンパク質のスペクトルに差異を引き起こすようである。なぜならこの原子は、上記タンパク質中のペプチド結合に埋め込まれているからである。一部はこの構造に基づいて、Q、H、K、もしくはRに由来する窒素は、上記モチーフの1位におけるNの代わりに使用され得る。
【0073】
上記金属結合部位および配位の幾何的配置を研究した。PRL−1が二価金属カチオンに結合するという知見は、興味深い。なぜなら、他のPTPaseによる特異的金属配位は報告されていなかったからである。PRL−1のC末端配列は、この酵素ファミリーの中でも独特であり、GGH様モチーフ(特定の場合においては、金属に結合し得るが、この場合では、結合しない)および上記新規なMAPタグモチーフの両方をコードする。上記GGH様モチーフの幾何的配置は、平面四角形である。濃縮したPRL−1サンプルで認められた徴的なさび色は、上記GGHペプチドの色と類似する。このことは、Ni配位が、平面四角形の幾何的配置において起こることを示唆する。本発明者らの研究から、NiがPRL−1のC末端において結合されることがさらに明らかになっている。金属配位は、PRL−1中の上記GGH様モチーフによって達成されない。なぜなら、上記おそらく必須のHisの変異は、結合に対してほとんど影響を有さないからである。両方のトリペプチドが平面四角形の配位を使用して結合するという事実にも拘わらず、各々についての化学現象は異なる。
【0074】
Ni精製WT PRL−1から得られたUV−Visスペクトルは、318nm、421nmおよび526nm付近にバンドを示す。平面四角形の幾何的配置において窒素配位子を介してNiを結合したペプチドおよび有機分子については、420nm付近の特徴的なd−dバンドが、文献中で頻繁に報告されている。Maroneyらは、最初から計算を行い、平面四角形の幾何的配置を使用して、Ni(II)およびCu(II)を配位する合成ペプチド模倣物のスペクトル割り当てを行った。1つのバンドを除いて、彼らの観察および計算は、PRL−1から観察されたスペクトルと非常に近く一致している。別個に、NiSODの還元した平面四角形形態を、密度汎関数理論を使用して分析した。その結果は、類似の関連する特徴を示す。これらシステムにおける上記バンドの位置およびそれらの相対強度の傾向は、上記PRL−1タンパク質スペクトルにおいて十分に保存されている。これらおよび他の研究に基づいて、526nmバンドは、Nixy−−>Nix2−y2転移から生じるようである。同様に、上記421nmバンドは、Niz2−−>Nix2−y2転移からくる可能性がある。強い上記318nmバンドは、上記配位子−金属電荷転移バンド(LMCT)を反映する。上記スペクトルの分解から、この吸収が、306nm、325nm、および372nmにおける3つのバンドが複合しているようであることが明らかにされている。他のガイドとしての構造を使用して、306nmにおける最も鋭く最も強いバンドが、硫黄原子からの転移に対応し、他の2つのバンドは、窒素供与またはおそらくより弱い硫黄供与から生じる可能性がある。
【0075】
一連のC末端変異体の中で、C170およびC171を含む変異のみが、これら波長でのシグナルを破壊した。最も顕著なことには、上記LMCTに対応する318nmエンベロープ(envelope)は、上記システイン変異スペクトル中に不足している。全体の吸収プロフィールに関して、上記システイン変異体の低い300nm領域における上記小さな非常に広い吸収は、より低い波長における高強度のバンド、または散乱効果からの人工産物であるようである。なぜなら、それは、上記318nmの成分バンドのうちの1つであると不正確に位置づけられているからである。Ni d軌道(421nm、526nm)内の転移に特徴があり得るバンドはまた、硫黄が酸素によって置換される場合には存在しない。元のバンドは存在せず、さらなるバンドは、C170もしくはC171のいずれかの変異の際に吸収スペクトル中に現れる。また、これら残基に関わるジスルフィド結合は、上記MSデータにおいて観察されない。よって、両方のシステインが、金属配位に直接関与するようである。本発明者らのICP−MSデータは、C170およびC171に由来する両方の硫黄原子が、堅いNi結合に必要とされることをさらに示す。なぜなら、いずれの硫黄原子も、単独では十分ではないようであるからである。
【0076】
PRL−1によって生成されるスペクトル特徴はまた、Niが、残っている2つの位置における窒素原子によって配位されることを示唆する。頻繁には、Hisに由来するイミダゾリウムのNもしくは骨格アミドのNは、金属(例えば、Zn、CuおよびNi)を配位することが観察される。平面四角形の幾何的配置を使用して、Niを結合するモデルペプチドおよびタンパク質において、上記繋がる窒素基のうちのいくつかは、脱プロトン化され、負に荷電した種が、上記金属を結合する。上記526nmにおける吸収バンドは、PRL−1が、脱プロトン化窒素を介してNiを配位することを示すのに対して、上記PRL−1スペクトルにおいて生じた421nm吸収は、単独でプロトン化された窒素による配位を反映する。PRL−1−H166Aを作製するための上記GGH様モチーフにおけるHisの変異は、このヒスチジンの側鎖が、Ni結合に実質的に影響を及ぼさないことを明らかにした。PRL−1は、H166に加えて、3つの他のHisをコードする。H23、H64およびH103は、上記配列の異なる領域に位置し、変性条件下で堅く結合したままである可能性は低い。上記金属を放出するために上記タンパク質を解こうとする試みは、高温で数時間にわたる高濃度の変性剤での激しい処理を必要とした。化学的変性および熱変性は、Niのゆっくりとした放出のみを可能にし、CDデータは、Niが放出される十分前にPRL−1が完全に解かれることを示すので、上記金属は、上記タンパク質の短いセグメントにおいてクラスター形成した原子によって配位されるようである。
【0077】
上記GGHタイプペプチドは、極めて堅い結合(Kd 約10−17〜10−18)を達成する。このことは、上記モチーフ内の脱プロトン化骨格アミド窒素によって一部達成される。PRL−1について観察される極めて堅い結合および窒素原子からNi連結への推定される寄与に鑑みれば、PRL−1に由来する骨格アミドは、金属配位に関与し得る。いずれかのシステインの非存在が、結合を破壊し、短縮が両方の側鎖および骨格アミドを除去するので、金属交換反応が、変異誘発を使用して、C170およびC171に隣り合うアミド窒素に関わるという仮説を試験することは、可能でない。構造研究は、さらなる配位原子を同定する最もよい方法であり、これら研究は、本発明者らの実験室で進行中である。
【0078】
(実施例7)
生理学的pHにおけるCu−NCCを、生理食塩水バッグによって囲まれたチューブの中に入れ、University of Kansas Medical CenterにあるHoglund Brain Imaging Centerにおいて、標準的MRIを使用して画像化した(図4)。図4において、中心にある明るいシグナルは、上記Cu−NCC錯体に対応するのに対して、あまり明るくないシグナルは、上記生理食塩水溶液に対応する。黒色は、上記チューブおよび空気に対応する。
【0079】
従って、本発明は、言及した目的および利点、ならびにそこに固有のものを得るために十分に適合される。本発明は、その趣旨もしくは本質的な特徴から逸脱することなく、他の特定の形態において具現化され得る。記載される実施形態は、単なる例示として全ての観点において考慮されるべきであり、限定として考慮されるべきではない。本発明の範囲は、従って、前述の説明によってではなく、添付の特許請求の範囲によって示される。特許請求の範囲の等価物の意味および範囲内に入る全ての変更は、特許請求の範囲の範囲内に包含されるべきである。本明細書で記載される全ての参考文献は、具体的参照によって本明細書に参考として援用される。
【技術分野】
【0001】
(関連する出願との相互参照)
本願は、米国仮特許出願第61/052,918号(2008年5月13日出願)に対する優先権を主張する。この仮特許出願は、参照により本願に援用される。
【0002】
(政府の利益の陳述)
本発明は合衆国国立衛生研究所の助成金(Grant No.P20 RR−17708)の下、開発された。合衆国政府は本発明に対し一定の権利を有し得る。
【背景技術】
【0003】
(背景)
以前に、金属を利用して、ポリペプチド(例えば、タンパク質もしくはタンパク質フラグメント)を組成物から抽出することは、一般的な技術であった。このような抽出物は、上記組成物内の他の物質に優先して、金属が特定のポリペプチドと錯体形成する能力に基づいた。2つの異なるペプチドベースの金属結合タグが、組成物からペプチドを分離するために使用され得ることは、公知である。しかし、これら結合タグは、アミノ酸の特定のタイプ、および本発明に関連して記載されるものとは実質的に異なる結合を達成するために利用されるアミノ酸の特定の数もしくは特定の配列を有する。例えば、例えば、これら2種の結合タグは、同じタイプの金属に結合しなくてもよいし、本発明と同じ幾何的配置(geometry)を使用して結合しなくてもよい。第1の結合タグに関するさらなる情報は、特許文献1(Haroonら)において得られ得、配列NXEQVSPを含むペプチドに関する。第2の結合タグに関する情報は、特許文献2(Arbogastら)において得られ得、そこで記載されるタンパク質全体であるようであるタグの配列に関する。
【0004】
タンパク質の組換え発現のために生物の遺伝物質においてコードされ得るタグは、タンパク質生成物の精製および同定のために広く利用されてきた。この最も顕著な例は、His−タグ技術であった。この技術は、固定化金属アフィニティークロマトグラフィー(IMAC)を使用して、全細胞からタグ化したタンパク質を効率的に単離する容易な手段を提供する。多くの他のペプチドベースのタグは、ペプチドタグを認識する抗体を使用して、細胞培養アッセイもしくは細胞溶解物において上記タグ化タンパク質を検出するために開発されてきた。これら技術は、インサイチュアッセイもしくはインビトロアッセイにおいて有用であるが、一般に、それらはインビボ分析には適用可能でない。ペプチドタグの利点は、上記タグが、目的のタンパク質を標識するためのさらなる化学工程の必要なくして、上記タンパク質に共有結合することである。
【0005】
MRI画像化は、生きている動物およびヒトにおいて構造的特徴を調べるための一般的な方法である。上記技術は安全でありかつ非侵襲性である。造影剤は、上記方法の感度を改善するために開発されてきており、それらは、上記MRI画像において認められる特徴を増強するために使用される。上記改善は、造影剤が、隣り合う分子(代表的には、水)からのシグナルを変化させる金属を含むことから、達成されてきた。最も一般に使用される薬剤は、Gd(III)をキレート化するが、他の金属は、対比を増強するために使用され得る。Gd含有造影剤は、腎性全身性線維症もしくは腎性線維化性皮膚症として公知の重篤な合併症が生じ得るので、腎機能が損傷した患者において使用できなかった。このような患者の必要性に対処するために、代替の画像化剤が必要とされる。金属をキレート化する化合物はまた、分子レベルでの詳細を検討するために、PET画像化およびSPECT画像化において使用される。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】米国特許第7,208,138号明細書
【特許文献2】米国特許出願公開第2004/0018974号明細書
【発明の概要】
【課題を解決するための手段】
【0007】
(要旨)
本開示は、一般に、トリペプチドモチーフおよびこのようなモチーフを使用するための方法に関する。これらペプチドは、金属に結合する能力を有し、このことは、これらペプチドを種々の適用に有用にする。特に、本開示のトリペプチドは、画像化、研究、化学療法、およびキレーション療法(chelation therapy)における適用を有する。
本発明は、例えば以下の項目を提供する。
(項目1)
配列XC1C2を有するトリペプチドを含む組成物であって、ここでXは、XC1C2が平面四角形配向もしくは四角錐配向またはその両方において金属を結合し得るような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、組成物。
(項目2)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目1に記載の組成物。
(項目3)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目1に記載の組成物。
(項目4)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目1に記載の組成物。
(項目5)
配列XC1C2を有するトリペプチドおよび金属を含む組成物であって、ここで該金属は、該トリペプチドと錯体化しており;そしてここでXは、トリペプチドおよび金属が平面四角形配向もしくは四角錐配向またはその両方を有する錯体を形成するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、組成物。
(項目6)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目3に記載の組成物。
(項目7)
前記金属は、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeからなる群より選択される、項目3に記載の組成物。
(項目8)
前記金属は、放射性同位体である、項目3に記載の組成物。
(項目9)
前記金属は、62Cuである、項目3に記載の組成物。
(項目10)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目3に記載の組成物。
(項目11)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目3に記載の組成物。
(項目12)
金属と、配列XC1C2を有するトリペプチドとを錯体化して、金属−XC1C2錯体を形成する工程を包含する方法であって;ここでXは、金属−XC1C2錯体が、平面四角形配向もしくは四角錐配向またはその両方を有するような任意のアミノ酸であり;そして
ここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される、方法。
(項目13)
Xは、アスパラギン、グルタミン、ヒスチジン、リジン、およびアルギニンから選択される、項目12に記載の組成物。
(項目14)
前記金属は、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeからなる群より選択される、項目12に記載の組成物。
(項目15)
前記金属は、放射性同位体である、項目12に記載の組成物。
(項目16)
前記金属は、62Cuである、項目12に記載の組成物。
(項目17)
前記配列XC1C2を有するトリペプチドを含むポリペプチドをさらに含む、項目12に記載の組成物。
(項目18)
前記配列XC1C2を有するトリペプチドを含む非ペプチド実体をさらに含む、項目12に記載の組成物。
(項目19)
前記金属は、生物学的サンプル中に存在する、項目12に記載の方法。
(項目20)
前記金属は、被験体において見いだされる、項目12に記載の方法。
(項目21)
前記金属−XC1C2錯体を検出する工程をさらに包含する、項目12に記載の方法。
(項目22)
前記金属−XC1C2錯体を画像化する工程をさらに包含する、項目12に記載の方法。
【0008】
特定の実施形態によれば、本開示は、配列XC1C2を有するトリペプチドを含む組成物を提供し;ここでXは、XC1C2が平面四角形配向もしくは四角錐配向またはその両方において金属を結合し得るような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0009】
他の実施形態によれば、本開示は、配列XC1C2を有するトリペプチドおよび金属を含む組成物を提供し;ここで上記金属は、上記トリペプチドと錯体化し;そしてここでXは、トリペプチドおよび金属が、平面四角形配向もしくは四角錐配向またはその両方を有する錯体を形成するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0010】
他の実施形態によれば、本開示は、金属と、配列XC1C2を有するトリペプチドとを錯体化して、金属−XC1C2錯体を形成する工程を包含する方法を提供し;ここでXは、金属−XC1C2錯体が、平面四角形配向もしくは四角錐配向またはその両方を有するような任意のアミノ酸であり;そしてここでC1およびC2は、同じであるかまたは異なっており;そしてここでC1およびC2は、システインおよびシステイン様非天然アミノ酸から個々に選択される。
【0011】
本発明の特徴および利点は、以下の実施形態の説明を読めば、当業者に容易に明らかである。
【0012】
(図面)
本開示のより完全な理解は、添付の図面とともに、以下の詳細な説明を参照することによって得られ得る。
【図面の簡単な説明】
【0013】
【図1】図1は、構造1 4N配位を有するGGH様金属錯体NGHを示す。構造2および構造3 いくつかの脱プロトン化骨格N原子によって上記金属の配位に影響を及ぼすGGH様配置を示す。
【図2】図2は、ヒトPRL酵素(PRl−1、PRL−2、およびPRL−3を含む)のC末端部分の配列アラインメントを示す。PRL−1のH166は、青で示されるNGHモチーフの一部である。PRL−1の残基C170およびC171は、赤で示されるCaaXモチーフ(ボックスで囲った)の一部である。上記NCCモチーフは、太字が付されている。
【図3】図3は、A. pH10において獲得したニッケル結合トリペプチドのESI−MS(ここで質量は、ニッケルが上記ペプチドに結合されていることを示す)、B.上記サンプルのpHを5に低下させた後のESI−MSスペクトル(ニッケルが、上記ペプチドから放出されたことを示す)を示す。
【図4】図4は、Cu−NCC磁気共鳴画像(MRI)を示す。
【図5】図5は、His−タグの切断および除去後に、Ni(II)(pH7.4)の存在下で精製した、精製WT PRL−1の写真を示す。写真は、その特徴的なさび色を示す。
【図6A】図6Aは、Ni2+イオンの存在下で精製したPRL−1改変体の電子吸収スペクトルを示す。各タンパク質改変体について3つの(2つの)スペクトルを800〜200nmまで記録し、平均した。全ての改変体についてのタンパク質濃縮物は、A280比較に基づいて、5mg/mLから10%以内までである。700〜300nmまでのスペクトルを示す。目に見える特徴は、318nmおよび421nmでの吸収極大である。挿入図。421nmおよび526nmにおいて吸収極大の可視化のために、より小さなスケールで、600〜400nmまでのスペクトルが示される。曲線は、以下のとおりである:PRL−1−WT、黒(一番上);PRL−1−H166A、赤(一番上から2番目);PRL−1−C170S−C171S、橙色(一番下);PRL−1−C170S、緑(一番下から2番目);PRL−1−C171S、青(真ん中)。
【図6B】図6Bは、PRL−1 WTの分解スペクトル(Resolved spectrum)。300〜700nmの間のNi精製PRL−1 WTタンパク質の上記UV−Visスペクトルの分解は、5つの曲線を使用するデータと適合する。その318nmのショルダーは、306nm、325nm、および372nmにおけるピークに分解される。上記分解された吸収ピーク(灰色破線)の和および生データ(黒実線)が、ともに示される。421nmおよび526nmにおけるピークもまた、存在する。
【0014】
特許および特許出願ファイルは、少なくとも1部のカラーで仕上げられた図面を含む。カラー図面付きのこの特許もしくは特許出願公開のコピーは、要求および必要な料金を支払えば、特許庁によって提供される。
【0015】
本開示は、種々の改変および代替の形態に影響されやすいが、特定の例示的実施形態は、図面に既に示されており、本明細書により詳細に記載される。しかし、特定の例示的実施形態の説明が、本発明を開示される特定の形態に限定すると意図されないが、一方で、この開示は、添付の特許請求の範囲によって規定されるような全ての改変および等価物を網羅することが理解されるべきである。
【発明を実施するための形態】
【0016】
(説明)
本発明は、一般に、短いペプチドモチーフおよびこのようなモチーフを使用するための方法に関する。これらペプチドは、選択的に金属に結合する能力を有し、このことは、これらペプチドを、種々の適用に有用にする。特に、本発明のペプチドは、画像化、研究、化学療法、およびキレーション療法における適用を有する。
【0017】
一般に、本開示は、選択された金属と強く結合する短い新規なペプチドモチーフ(MAPタグといわれる)に関する。よって、これらMAPタグは、とりわけ、上記選択された金属を、組成物から抽出するために使用され得る。本開示のMAPタグは、3アミノ酸長であり、N末端、C末端、もしくはその間の任意の位置において、より長いポリペプチドおよびタンパク質の中に含まれ得る。特定の実施形態において、しかし、上記MAPタグが、金属と結合するための上記MAPタグを表す、ポリペプチドもしくはタンパク質の配置に存在する(例えば、外部ループに存在する)ことは、有利であり得る。上記MAPタグはまた、非ペプチド実体に結合され得る。さらに、1種より多いMAPタグは、特定の分子上に存在し得る。
【0018】
多くのタンパク質が金属に結合することは公知である。しかし、このような結合は、代表的には、密な結合を達成し、次いで、開いた際に、容易に上記金属を放出するために、折りたたまれたタンパク質構造内で不連続なアミノ酸配列を利用する。他方で、MAPタグによる金属結合は、非常に密接して近くにある原子を使用して達成され、よって、極端な条件が、上記金属を放出するために必要とされる。上記タンパク質の熱変性および化学的変性は、上記金属のゆっくりとした放出を可能にする。例えば、極端な条件の使用(例えば、沸点温度、強酸)は、一定期間にわたって(例えば、数時間から長時間)上記金属をゆっくりと放出することをもたらし得る。
【0019】
(A.化学的構造)
特定の実施形態において、本開示は、新規なペプチドベースのタグもしくは金属の結合剤(MAPタグ)を提供する。上記MAPタグは、ポリペプチドもしくはタンパク質中に高親和性金属結合部位を直接コードするために使用され得る。本開示のMAPタグは、平面四角形および/もしくは四角錐の幾何的配置において金属に結合し得るそれらの能力によって特徴付けられる。
【0020】
(1.ペプチド配列)
本開示のMAPタグは、一般に、金属に結合し得る、少なくとも3個連続するアミノ酸残基を含む。本開示のMAPタグは、一般に、XC1C2によって表される配列を有し、ここでC1およびC2は、同じであってもよいし、異なっていてもよく、システイン、もしくはシステイン様非天然アミノ酸(例えば、硫黄含有α−アミノ酸もしくはβ−アミノ酸)であり得、そしてここでXは、上記形成されるペプチドタグが、平面四角形/四角錐の幾何的配置において金属に結合し得る限りにおいて、別の天然アミノ酸もしくは非天然アミノ酸、またはアミノ酸アナログであり得る。本明細書において使用される場合、L鏡像異性アミノ酸の略語は、従来どおりであり、以下のとおりである:
【0021】
【表1】
いくつかの実施形態において、Xは、アスパラギン(N)、グルタミン(Q)。ヒスチジン(H)、リジン(K)、もしくはアルギニン(R)であり得る。
【0022】
アミノ酸配列が、一連の三文字アミノ酸略語もしくは一文字アミノ酸略語として表される場合、標準的な使用および慣例に従って、左側がアミノ末端方向であり、右側がカルボキシ末端方向であることが理解される。
【0023】
本開示のMAPタグは、任意の組換え技術システムを使用して、発現のための遺伝子もしくはヌクレオチド配列と一続きで(in line)コードされ得る。さらに、上記タグは、ペプチドもしくはタンパク質の生成のための任意の合成法もしくは生合成法を使用して、ペプチドもしくはタンパク質へと組み込まれ得る。適用において、上記MAPタグは、金属と自発的に反応して、ペプチド−金属錯体を形成する。このようなペプチド−金属錯体は、溶液中で、または金属交換反応(transmetallation)もしくは任意の他のプロセスを介して、形成し得る。
【0024】
上記MAPタグは、単独で使用され得るか、あるいはN末端側、C末端側に結合され得るか、またはポリペプチド鎖内の可撓性リンカーもしくはループへと挿入され得るタグとして、使用され得る。上記MAPタグは、金属配位に関与する側鎖以外の任意の位置で改変され得る。よって、上記MAPタグは、Z−XC1C2−Z1のような配列を含み得、Zは、任意のアミノ酸もしくは任意のアミノ酸配列であり得、Z1は、Zに等しいもしくは等しくない、任意のアミノ酸またはアミノ酸配列であり得る。上記アミノ酸は、上記表1と同様に示される。非天然のおよびアミノ酸のアナログは、上記表中には列挙されていないが、それらは、Z、Z1、および/もしくはXに組み込まれ得る。
【0025】
特定の実施形態において、上記MAPタグは、別の分子に結合され得る。例えば、上記MAPタグは、炭水化物(例えば、ヒアルロン酸)のような非ペプチド実体に結合され得る。上記結合は、共有結合であってもよいし、リンカーを介して行われていてもよい。
【0026】
いくつかの実施形態において、上記MAPタグは、以下のような配列を含み得る:NC1C2;Z−NC1C2−Z1;Z−NC1C2;NC1C2−Z1;QC1C2;Z−QC1C2−Z1;Z−QC1C2;QC1C2−Z1;HC1C2;Z−HC1C2−Z1;Z−HC1C2;HC1C2−Z1;KC1C2;Z−KC1C2−Z1;Z−KC1C2;KC1C2−Z1;RC1C2;Z−RC1C2−Z1;Z−RC1C2;もしくはRC1C2−Z1。上記のように、Zは、任意のアミノ酸もしくは任意のアミノ酸配列であり得、Z1は、またはZに等しいかもしくは等しくない、任意のアミノ酸もしくはアミノ酸配列であり得る。上記アミノ酸は、表1と同じように表される。非天然のおよびアミノ酸のアナログは、上記表中には列挙されていないが、それらは、化合物ZおよびZ1に組み込まれ得る。
【0027】
一実施形態において、上記MAPタグは、以下の構造式を有するNCCを含む:
【0028】
【化1】
特定の実施形態において、本開示のMAPタグは、金属とのMAPタグ錯体化前、もしくは金属との錯体化後のいずれかに、上記MAPタグの標的化送達を提供する遺伝子もしくはヌクレオチド配列と一続きでコードされ得る。このことは、遺伝子、ペプチド、もしくは標的化に有用であることが公知の他のモチーフを使用して、達成され得る。例えば、MAPタグは、抗体、増殖因子、ペプチドなどと一緒に組み込まれ得る。
【0029】
(2.金属結合)
特定の実施形態において、本開示のMAPタグは、単独で、またはポリペプチドもしくはタンパク質に組み込まれる場合、金属と錯体化して、平面四角形/四角錐の幾何的配置を有するMAPタグ−金属錯体を形成し得る。上記金属は、上記MAPタグと、2N:2S配位を介して錯体化し得る。上記MAPタグはまた、当業者によって認識される適切な条件下で、平面四角形の幾何的配置において多くの金属に結合し得る。一般に、本開示のMAPタグは、IUPAC Groupを参照すると:3族金属(例えば、Y);5族金属(例えば、VおよびU);6族金属(例えば、Cr、Mo、W);7族金属(例えば、Mn、Tc、Re);8族金属(例えば、FeおよびRu);9族金属(例えば、Co、Rh、Ir);10族金属(例えば、Ni、Pd、Pt);11族金属(例えば、Cu、Ag、Au);12族金属(例えば、Zn、Cd、Hg);13族金属(例えば、Al、Ga、In、Tl);14族金属(例えば、SnおよびPb);ならびに15族金属(例えば、Bi)を結合し得る。特定の実施形態において、上記MAPタグは、Zn、Ni、Cu、Pt、Pd、Au、Ag、Pb、およびFeと結合しかつMAPタグ−金属錯体を形成し得る。
【0030】
上記のように、上記金属を結合すると、上記MAPタグは、平面四角形の幾何的配置を有する。本開示はまた、種々の平面四角形配置にある、MAPタグ金属配位を企図する。例えば、上記MAPタグがNCCでありかつ上記金属が上記のようにMとして表される任意の金属である場合、上記MAPタグは、式1によって表される構造式を有し得る:
【0031】
【化2】
他の構造的配置がまた、企図される。例えば、上記MAPタグがNC1C2でありかつC1が非天然アミノ酸(例えば、(S)−2−アミノ−2−メルカプト酢酸)でありかつC2がシステインである場合、上記MAPタグの構造式は、式2として表され得、ここでMは、上記のように任意の金属である:
【0032】
【化3】
別の例において、上記MAPタグは、NC1C2であり、C2は、非天然アミノ酸(例えば、(S)−2−アミノ−4−メルカプト酪酸であり、上記MAPタグの構造式は、式3として表され得、ここでMは、上記のように任意の金属である:
【0033】
【化4】
別の例において、上記MAPタグは、NC1C2であり、C2は、βアミノ酸(例えば、3−アミノ−2−メルカプトプロピオン酸であり、上記MAPタグの構造式は、式4として表され得、ここでMは、上記のように任意の金属である:
【0034】
【化5】
別の例において、上記MAPタグは、RCCであり、上記MAPタグの構造式は、式5として表され得、ここでMは、上記のように任意の金属である:
【0035】
【化6】
別の例において、上記MAPタグは、QCCであり、上記MAPタグの構造式は、式6として表され得、ここでMは、上記のように任意の金属である:
【0036】
【化7】
(B.上記MAPタグの適用)
(1.金属抽出のためのMAPタグ)
上記のように、MAPタグは、高親和性で金属を結合し得る。MAPタグはまた、液体から固体までの範囲に及ぶ種々の組成物から金属を抽出し得る。結論として、MAPタグが、配位を共有するのではなく上記金属を抽出する能力は、以下で依り詳細に記載されるように、上記タグを、特定の金属を別の組成物から分離することにおける使用に影響を受けやすくする。上記MAPタグは、上記金属と錯体化し、次いで、上記金属を組成物中の成分(例えば、キレート化剤(例えば、EDTA)、または例えば、IDAもしくはNTAと結合体化した固体支持体)から上記金属を抽出もしくは除去することによって、上記組成物から金属イオンを封鎖する。よって、上記MAPタグは、金属抽出ペプチド(MAP)タグである。
【0037】
さらに、少なくとも一部は、MAPタグが金属に選択的に結合する能力に起因して、本開示の実施形態は、キレーション療法に有用であり得る。従って、MAPタグは、生物学的に毒性の金属(例えば、鉛)を封鎖するために使用され得る。
【0038】
同様に、MAPタグはまた、金属を水、他の水溶液、もしくは身体の外の任意の組成物から封鎖することにおいて有用であり得る。上記MAPタグ自体は、一般に、多くの溶媒中で可溶性である。しかし、特定の実施形態において、上記MAPタグ−金属錯体は、多くの有機溶媒中では溶解性が乏しく、いったん金属結合が起こると、沈殿し得る。このことは、有機溶媒からの抽出の間の効率的な分離を可能にする。
【0039】
(2.画像化適用)
特定の実施形態において、本開示は、MAPタグを使用する、画像化のための方法を提供する。このような方法は、治療的画像化および医療的画像化、または他の画像化適用のために使用され得る。上記MAPタグと錯体化するように選択される金属は、上記特定の適用および画像化技術に依存する。例えば、常磁性金属は、画像化技術(例えば、MRI、NMRおよびEPR)との使用に選択され得る一方で、金は、電子顕微鏡のような技術のために選択され得る。その一方で亜鉛は、蛍光のような適用のために選択され得る。どの金属が所定の適用および画像化技術に適しているかは、当業者によく知られている。
【0040】
一実施形態において、MAPタグは、画像化技術において使用するための試薬を調製するために利用される。すなわち、MAPタグは、金属に結合し、続いて、画像化のために使用され得る(例えば、上記金属が画像化のために使用され得るように、被験体の身体において使用され得る)錯体を提供する。適切な画像化技術の例としては、単独でもしくは組み合わせにおいて、コンピューター断層撮影法(CT)、磁気共鳴画像法(MRI)、超音波、陽電子放射断層撮影法(PET)、単一光子放出型コンピューター断層撮影法(SPECT)、核磁気共鳴法(NMR)、電子常磁性共鳴(EPR)、電子顕微鏡、蛍光画像法などが挙げられる。
【0041】
別の実施形態において、MAPタグは、被験体の一部に局在するポリペプチドもしくはタンパク質、または非タンパク質実体へと組み込まれ得る。ここでMAPタグ−金属錯体の金属部分は、局在部位(例えば、組織もしくは腫瘍)を画像化する能力を提供する。例えば、上記MAPタグ−金属錯体は、上記MAPタグ化分子の標的化部分に対するレセプターおよび/もしくは結合パートナーを発現する組織もしくは他の構造を同定するために使用され得る。上記金属を可視化し得る画像化技術は、このような特徴の同定のために使用され得る。
【0042】
例えば、MRI造影剤は、隣り合う原子の緩和を変化させて、上記造影剤が存在する領域と、存在しない領域との間で検出されるシグナルの差異を発生させる、代表的には、金属中心を含む常磁性種である。MAPタグは、Cuと錯体化される場合には、常磁性であり得る。従って、MAPタグ−金属錯体は、MRI造影剤として使用するための能力を有する。大部分の造影剤は、顕微鏡レベルで標的化されないが、身体もしくは組織中の肉眼的構造および欠損を明らかにするために使用される。適用のこのタイプにおいて、上記造影剤の濃度は、非常に高い。造影剤を、選択された組織もしくは顕微鏡的構造に標的化することは、細胞レベルでの異常の同定を可能にする点で有益である。例えば、上記MAPタグは、特定のレセプターを発現する癌細胞を認識する抗体上のタグとして発現され得る。標的化された送達は、より低い濃度の造影剤が投与されることを可能にする。このことは、とりわけ、毒性および他の副作用を最小にする助けになり得る。
【0043】
62Cuが、身体内の分子的な詳細を画像化し得る、PETおよびSPECT画像化において使用される放射活性同位体であることは公知である。よって、MAPタグは、Cu(もしくは別の金属)の適切な放射性同位体を添加され得、これら画像化技術でのトレーサーとして使用され得る。同様に、上記タグを、任意のタンパク質もしくはペプチド、または細胞の部分セットを結合する他の分子に融合することによって、標的化された画像化が、達成され得る。
【0044】
上記MAPタグは、特定の金属に対して高い親和性を有する短い配列であるので、とりわけ有利である。従って、発現されたタンパク質もしくは他の分子においてMAPタグをコードすることによって、生物内の特定の細胞タイプへの標的化された送達は、可能である。さらに、細胞の部分セットを標的化することは、画像化に必要とされる造影剤の量の減少を可能にする。
【0045】
(3.化学療法)
本発明はまた、化学療法を補助するために使用され得る。例えば、MAPタグは、白金および/もしくは放射性核種との錯体を形成し得、特定の細胞もしくは細胞タイプへの標的化された送達を提供するために使用され得る。
【0046】
本発明のよりよい理解を促進するために、以下の特定の実施形態の実施例が提供される。以下の実施例が、本発明の範囲全体を限定するようにも、規定するようにも、決して解釈されるべきではない。
【実施例】
【0047】
(実施例1)
変異誘発技術を、選択された金属と強い結合親和性を有する上記ペプチドモチーフを同定するために使用した。簡潔には、ヒトPRL−1遺伝子を、pET−30 Xa LICベクター(Novagen)にクローニングし、PCRベースのQuikChange法(Stratagene)を使用して変異させた。C170S、C171S、およびH166A変異体を生成するプライマー(5’から3’)は、それぞれ、
【0048】
【数1】
および
【0049】
【数2】
(Integrated DNA Technologies,Coralville,IA)であった。大文字は、変異した塩基を示す。PCR反応物をDpnI(Promega)で、1.5時間にわたって37℃において処理し、NovaBlue GigaSingles Competent細胞(Novagen)に直接形質転換し、30mg/mL カナマイシン選択でLB上にまいた。個々のコロニーを、選択的LB中で37℃において一晩増殖させた。得られたDNAを、Wizard Plus MiniPrep System(Promega)を使用して精製した。全ての変異を、DNA色素−ターミネーター配列決定によって確認した(Northwoods DNA,Inc.,Bemidji,MN)。上記二重変異体C170S/C171Sを、第2回の変異誘発によって生成し、同様に確認した。
【0050】
PRL−1および関連する変異遺伝子を有するベクターを、BL21(DE3)細胞(Novagen)へと形質転換し、選択的LB上で37℃において一晩増殖させた。選択されたコロニーを、選択的M9ZBブロス中で、一晩37℃において増殖させた。次いで、一晩の培養物を、500mLの非標識最小培地に移し、0.6〜0.8の間のOD550において、1mM IPTGで誘導した。NMR実験のために発現させたタンパク質は、15N−塩化アンモニウムを含む最小培地中で増殖させた。上記組換えタンパク質の過剰発現を、SDS−PAGEによって確認した。細胞を、4400×gでの遠心分離によってペレット化し、使用するまで−80℃で保存した。
【0051】
一連のPRL−1変異体を、金属配位に関与する残基を同定するために試験した。GGH様モチーフを含むモデルペプチドは、極めて高い親和性でNi(II)およびCu(II)を配位することを示した。このモチーフにおいて、2つのGly位置のうちのいずれかにおける置換が、ごくわずかに結合を低下させるのに対して、Hisの変異は、結合を破壊する。PRL−1は、NGH、GGH様コンセンサスモチーフを、そのC末端付近においてコードする(図2)。従って、PRL−1−H166Aを、PRL−1に含まれる上記NGH配列が、金属結合を担うという仮説を試験するために作製した。PRL−1およびPRL−1−H166Aを、それぞれ、約120μMおよび60μMのNiを結合したICP−MSによって、並行して分析した。吸収分光法によって測定した場合に、わずか20%のシグナルの低下が、H166Aについて認められた。タンパク質シグナルが、等しい程度にまで減少され、可視領域のシグナルの損失が、タンパク質不安定性に起因することはまた、吸収スペクトルから明らかである。タンパク質不安定性は、上記2つの検出法における不均衡が原因のようである。なぜなら、ICP−MSは、分析する前に、実質的により多くのサンプル操作を要するからである。上記ヒスチジン変異体は、堅い結合を保持したままであったので、C170および/もしくはC171における変異を、チオールがNi配位にしばしば関与することから、作製した。個々のシステイン変異体、ならびに二重変異体におけるNiレベルは、野生型タンパク質と比較して、大いに低下した。このことは、両方のCys残基が、金属配位に影響を及ぼすことを示す。C170S変異体においても、上記C170S−C171S二重変異体においてもNiが検出されなかったが、約17μMの小さなシグナルが、PRL−1−C171Sについて認められた。
【0052】
(実施例2)
タンパク質精製技術を使用して、上記タンパク質−金属錯体を精製した。簡潔には、細胞ペレットを、30mL 緩衝液A(100mM NaCl、50mM Tris−HCl、10mM イミダゾール、pH7.4、Ar散布した)中に再懸濁し、15,000psiにおいて、フレンチプレッシャーセル(ThermoElectron)を使用して溶解した。緩衝液のアルゴン散布を行って、酸素を置換した。次いで、サンプルを、21,000×gにおいて遠心分離し、その上清を、0.2μm ナイロンフィルタを通して濾過した。(His)6−タグ化PRL−1の精製を、流速1mL/分において、Akta Explorer精製システムを使用して行った。サンプルを、金属添加した5ml HiTrap Chelating Column(GE Healthcare)に載せ、載せた後に、5×カラム容積ですすいだ。溶出を、13×カラム容積かけて60% 緩衝液B(100mM NaCl、50mM Tris−HCl、500mM イミダゾール、pH7.4、アルゴン散布ありもしくはなし)へ、3×カラム容積かけて100% Bへの直線勾配を使用して達成し、および4×カラム容積かけて100% Bにおいて維持した。溶出を、280nmでの吸収によってモニターし、画分を、SDS−PAGEを使用して純度について試験した。適切な画分をプールし、Ar散布したもしくはAr散布していない100mM NaCl、50mM Tris−HCl、pH7.4に対して透析した。上記(His)6−タグを、第Xa因子プロテアーゼ(Promega)を、室温において一晩使用して、上記標的タンパク質から切断した。XarrestTMアガロース(Novagen)を使用して、プロテアーゼを除去し、上記カラムに再度載せて上記タグおよび任意の非切断タンパク質を除去する前に0.2μm ナイロンフィルタで濾過した。上記タンパク質を、透析緩衝液中の上記カラムに載せ、最初のフロースルーを集めて、上記タンパク質サンプルを作製した。サンプルを濃縮し、(Amicon Ultra)10kDa MWCO遠心分離フィルタを使用して、Ar散布100mM NaCl、50mM TrisCl、pH7.4中で、少なくとも106倍交換した。上記サンプルの純度および変異体確認を、それぞれ、SDS−PAGEおよびESI−MSによって確認した。カラムを、1M NaOHで処理し、2×カラム容積の100mM EDTAでストリッピングし、Ar散布した100mM NiSO4、100mM CuSO4、100mM CuCl2もしくは100mM ZnCl2を、各使用の直前に再度充填した。
【0053】
図5は、上記His−タグの切断および除去後の精製PRL−1を示す。上記his−タグに結合した金属は、八面体の幾何的配置を有し、これは、青緑色であり、このような配位事象に対応する吸収は、スペクトルのうちの700〜800nm領域において現れる。Ni(II),pH7.4の存在下で精製したWT PRL−1の写真は、その特徴的なさび色を示す。上記精製した野生型PRL−1タンパク質は、予測外のさび色を有する(図5)。このことは、芳香族残基(すなわち、TrpもしくはTyr)の金属錯体化もしくは酸化から生じ得る。
【0054】
芳香族残基の酸化は、上記タンパク質のインタクトなおよびトリプシン消化研究からの質量分析データに基づいて容易に排除された。さらに、本発明者らは、上記さび色のタンパク質が、高速遠心分離の間にチューブの底で濃縮されるので、上記タンパク質の密度が、予測されるより高いことを観察した。金属配位が上記タンパク質錯体の密度を増加させるので、本発明者らは、上記発色団が、上記タンパク質との金属錯体の形成によって生成される可能性をさらに調べた。金属キレート化クロマトグラフィーを使用して、PRL−1を精製したので、金属イオンのもっとも可能性のある供給源は、上記IMAC樹脂に由来するNiであった。
【0055】
組換えWT PRL−1を含む細胞溶解物が、等しく分けて、Ni2+充填した、Cu2+充填したもしくはZn2+充填したIMAC樹脂を使用して並行して(side−by−side)精製した場合、上記タンパク質は、上記キレート化樹脂に結合した金属を抽出する。金属交換反応の速度は、ニッケルで最も速く、続いて、銅であった。亜鉛は、全てのサンプルにおいてある程度存在し、このことは、上記タンパク質が亜鉛をインビボで拾い上げることを示す。亜鉛が上記カラムに結合される場合、亜鉛のみが、上記タンパク質中で認められる。Apo PRL−1は、可溶性が乏しく、金属の損失が沈殿をもたらす。誘導結合型プラズマ質量分析法(ICP−MS)によって試験した場合に各精製からの可溶性物質は、結合した金属の等しい割合を有する。
【0056】
(実施例3)
ICP−MSを使用して、上記結合した金属を特徴付けした。簡潔には、精製タンパク質サンプルを、PFA微小遠心分離管(Savillex,#s 7240、7241)において濃硝酸中、68±1℃で16.5±0.7時間にわたって消化した。消化前のタンパク質定量に基づいて、次いで、上記サンプルを、20mM 炭酸水素アンモニウム(pH8.2)中で1.5μMへと希釈し、0.2μm ナイロンフィルタを通して濾過し、微小同軸ネブライザ(microconcentric nebulizer)を装備したVG Elemental VGII+XS Inductively−Coupled Plasma−Mass Spectrometerへと注入した。最小で、2つの別個に調製したサンプルを、各タンパク質改変体について分析した。各サンプルを2回スキャンし、標準物質を、各サンプルスキャンの前、その間、およびその後に走らせた。ドリフトを、サンプルの注入と標準物質の注入との両方のスキャンの間の洗浄している間にモニターした。
【0057】
上記発色団が、荷電したIMACマトリクスからの金属のタンパク質抽出から生じるという本発明者らの仮説を調査するために、サンプルを、半定量的分析のためにICP−MSに供した。バックグラウンド金属を除去するために激しい洗浄を使用したので、この方法から得られた値は、結合した金属量より本質的に小さく見積もられるが、上記タンパク質に対して異なる金属の相対的親和性を評価するために使用され得る。さらに、上記細胞における発現の間にPRL−1に結合してしまった可能性のある内因性の金属リガンドを同定するために、ICP−MSを使用して、周期表中の最初の2列の遷移金属、および他の一般的に生物学的に関連する金属の存在を探した。顕著なことには、上記データは、Znおよび特定の金属(例えば、Ni)にのみ結合したPRL−1が、精製の間に上記IMACカラムで使用されたことを示した。16時間にわたって70℃において硝酸中で消化した、平均して500μM Ni精製タンパク質サンプルは、約180μM Niを含むと決定され、これは、1:2.7+/−0.7のNi 対 タンパク質化学量論を生じた。Znはまた、約60μMにおいて上記Ni精製サンプル中で検出された。別個に、Zn−IMACを使用するPRL−1の精製は、Znが、270μMにおいて多い量で存在する唯一の金属であり[1 Zn:1.8 タンパク質]、これらタンパク質サンプルが、Niのわずかなバックグラウンドレベルを含むことを示した。Ni精製PRL−1およびZn精製PRL−1の両方のICP−MSは、顕著な量で他の金属を示さず、このことは、亜鉛が、インビボで配位され、上記ニッケルのクロマトグラフィー精製の間に、Ni2+によって部分的に置換されたことを示唆する。ICP−MS分析は、Ar流中で行われ、Arの最も量の多い同位体がCaと同じ質量を有するので、Ca結合は、一次質量シグナル(primary mass signal)からは決定できない。にも関わらず、遙かに多い量のCaが、Znと比較すると細胞培養培地中に存在し、それでもなお上記ICP−MSシグナルが、主に亜鉛に対応するので、上記タンパク質は、カルシウムに結合しないようである。上記ICP−MSデータは、PRL−1がNi−IMACを使用して精製される場合、Znが評価できる程度の量で存在することを示す。このことは、Zn結合が細胞において存在することを示唆する。
【0058】
(実施例4)
UV−Vis吸収スペクトルを、上記PRL−1タンパク質の濃度を測定するために使用した。簡潔には、精製PRL−1アナログの濃度を、280nmにおいて測定し、19420M−1cm−1の吸光係数を使用して計算した。高い200nm領域での考えられる金属−タンパク質吸収干渉が原因で、上記UV−Vis定量法の有効性を、Bradford法によって確認した。10mg/mL、5mg/mL、2.5mg/mL、および1.25mg/mLの濃度のPRL−1アナログのスペクトルを、800〜200nmまで、Cary 100 UV−Vis分光光度計で集めた。上記分析における散乱効果を較正するために、PRL−1−WTスペクトルを、上記PRL−1−C170S−C171Sスペクトルを差し引くことによって調整した。なぜなら、上記PRL−1−C170S−C171Sスペクトルは、金属を結合しないからである。得られたスペクトルを分解し、ピーク位置を、GRAMS/AI 7.00ソフトウェアを使用して評価した。3つの方法は、金属置換を目的とした。第1において、PRL−1−WT(例えば、野生型)サンプルを、20mM β−メルカプトエタノール(BME)を使用して還元し、イミダゾールを、10mMの濃度になるように添加した。0.5mM タンパク質サンプルに対して、これは、BMEの400倍溶液過剰に対応して、上記2つのC末端システインチオールと競合し、イミダゾールの200倍溶液過剰に対応して、単一のC末端ヒスチジンイミダゾール側鎖と競合する。新たなベースラインをとり、スペクトルを再計算した。第2の方法において、ベースラインを、100μM EDTAを含む緩衝液について再計算し、100μM EDTA中で1000万倍交換した上記PRL−1−WTサンプルに対するスペクトルを、記録した。最後に、10mg/mL タンパク質サンプルを、5.6M グアニジウム塩酸塩中で変性させ、90℃へと加熱し、新たなベースラインをとり、そのスペクトルを、0時間、2時間、4時間、および6時間で集めた。
【0059】
よって、PRL−1による金属配位の特徴を、UV−Vis吸収分光法で決定した。PRL−1およびその変異体が使用した上記金属配位幾何的配置を、800〜200nmにおいてUV−Vis吸収分光法を使用して分析した。PRL−1の吸収スペクトルは、280nmおよび220nmにおいてピークを生成した。このことは、上記タンパク質内の芳香族およびペプチド結合から生じ、上記Bradfordアッセイを使用して決定される濃度とよく一致する。金属含有化合物は、可視範囲においていくつかの波長でしばしば吸収し、上記Ni精製およびCu精製のPRL−1タンパク質はまた、可視範囲においてピークを表し、これは、上記サンプルのさび色に対応する。PRL−1 WTは、318nmにおいて最大(非常に大きな280バンドのショルダーとして明らか)、421nmにおいてより広いシグナル、および526nmにおいて非常に広いピークを示す(図6A)。Cu精製PRL−1は、Ni精製PRl−1と同様のスペクトルプロフィールを生じる(データは示さず);しかし、小さい青色のシフトは、上記個々のスペクトルピークに対して観察され、このことは、CuをNiの代わりに使用した場合に予測される。予測されるように、Znを充填した樹脂を使用して上記タンパク質を精製した場合には、吸収も目に見える色も認められなかった。このことは、Zn(II)が完結したd殻を有し、よって、可視スペクトルにおいて電子移動を示すはずがないという事実に起因する。
【0060】
上記WTタンパク質と比較して、上記H166A改変体は、同じ吸収プロフィールを示したが、上記ピークは、上記ICP−MS結果と一致して、ある程度強度が低下した。PRL−1−WTおよびPRL−1−H166Aのスペクトルは、可視領域において上記システイン変異体のものとは異なる。等しい濃度において、システイン170および/もしくはシステイン171におけるセリンでの個々の置換は、421nmおよび526nmでのシグナルの完全な損失を示す。これら変異タンパク質を、10mg/mL(0.5mM)と同程度の濃度において試験したところ、特異的吸収バンドは検出されなかった。上記WTからのシグナルは、0.1mg/mLにおいて明らかであり、このことは、Niの結合が、各変異によって、少なくとも100倍低下することを示す。
【0061】
較正したWTタンパク質の上記UV−Visスペクトルのデコンボルーションを、300nm〜700nmの間で行った。すると、318nmピークが、それぞれ、306nm、325nm、および372nmにおいて、全ての複合ピーク吸収に対して54%、38%および7%寄与する3つの別個の吸収バンドから構成されることが明らかになる(図6B)。上記PRL−1−H166Aの吸収プロフィールは、上記野生型タンパク質スペクトルと非常に似ている。残基166における上記イミダゾール基の非存在は、吸収極大をシフトもせず、318nm吸収の成分バンドに顕著に寄与もしない。このことは、上記NGHモチーフ内のヒスチジンに由来するイミダゾールが、Niを直接配位しないことをさらに示す。
【0062】
PRL−1におけるNi結合は、PRL−1の活性部位におけるジスルフィド結合の還元によって影響を受けない。20mM BMEの添加は、上記スペクトル吸収プロフィールのUVもしくは可視領域を変化させず、このことは、金属結合が、上記タンパク質の還元および上記還元剤の存在によって大きく影響を受けないことを示す。上記還元型スペクトルを、上記酸化型から差し引くと、上記還元型WTサンプルと、上記酸化型WTサンプルとの間には、金属配位に有意差がないことが確認される。Hisからの小さな貢献がなされるか否かを調べるために、さらなるイミダゾールを、上記溶液に添加して、一時的な会合のシグナル強度を増大させた。10mM イミダゾールの添加は、上記吸収プロフィールに対して影響がなかった。このことは、上記Niへの接近が制限されていることを示唆する。さらに、上記Ni精製サンプルが、イミダゾールの存在下で還元された場合に、スペクトル変化は認められなかった。Ni精製WTサンプルの、100μM EDTAを含む緩衝液への1000万倍の交換は、上記スペクトルに対してほとんど影響がなく、このサンプルに対する吸光度値は、上記WT吸光度の誤差範囲内であった。7.4から8.5へとpHを上げると、モル吸収係数(ε)を約6倍増大させるはずであるが、興味深いことには、pHが上昇した場合に、εの変化は認められなかった。このことは、上記結合したNiが、PRL−1に接近できない溶媒であることを示唆し得る。しかし、6.5へと上記pHを下げると、目に見えるピークの強度は低下する(データは示さず)。
【0063】
Ni結合は、pH7.4において不可逆性になるようである。なぜなら、Ni精製PRL−1に由来するNiの移動が、攻撃的な手段を要するからである。Ni精製PRL−1を、5.6M グアニジウム塩酸塩で90℃において処理した場合に、318nm、421nmおよび526nmにおける上記ピークは、6時間かけて徐々に消失するに過ぎない(データは示さず)。750nmにおけるシグナルの付随した出現は、6時間かけて観察された。このことは、Niが、上記タンパク質から放出され、八面体の幾何的配置において水によって配意されたことを示す。図6Aは、Ni2+イオンの存在下で精製したPRL−1改変体の電子吸収スペクトルを示す。各タンパク質改変体に対する3つのスペクトルを、800〜200nmで記録し、平均した。全ての改変体についてのタンパク質濃度は、A280比較に基づいて、5mg/mLから10%以内までである。700〜300nmのスペクトルが示される。目に見える特徴は、318nmおよび421nmにおける吸収極大である。図6Aにおいて、曲線は、以下のとおりである:PRL−1−WT、黒色;PRL−1−H166A、赤色;PRL−1−C170S−C171S、橙色;PRL−1−C170S、緑色;PRL−1−C171S、青色。
【0064】
図6Bは、PRL−1 WTの分解したスペクトルを示す。300〜700nmの間でのNi精製PRL−1 WTタンパク質の上記UV−Visスペクトルの分解は、5つの曲線を使用した上記データと適合する。上記318nmのショルダーは、306nm、325nm、および372nmにおけるピークに分解される。上記分解された吸収ピークの和(灰色破線)および生データ(黒色実線)が、両方示される。421nmおよび526nmにおけるピークも、存在する。
【0065】
図6Bの挿入図は、600〜400nmのスペクトルを示し、421nmおよび526nmにおける吸収極大の可視化のために拡大スケールで示される。
【0066】
(実施例5)
エレクトロスプレーイオン化質量分析法(ESI−MS)を使用して、上記サンプルを分析した。簡潔には、ESIスペクトルを、MSモードで操作したQ−Tof−2(Micromass Ltd,Manchester UK) Hybrid Mass Spectrometerで獲得し、感度について8000分解(FWHH)に離調した(de−tuned)フライトアナライザーの時間とともにデータを獲得した。全タンパク質ESIスペクトルについては、精製タンパク質サンプルを、未処理(「酸化」)20mM 炭酸水素アンモニウム(pH8.2)中で1μg/μLに希釈し、室温でインキュベートした。精製後1日目、4日目、および7日目の時点で採取し、使用する準備が出来るまで−80℃で凍結した。サンプルを、短い(3cm×1mm内径)逆相(RP)HPLCカラム(Hamilton PRP1,Reno,NV)で脱塩した。サンプルを、タンパク質(5μg)を含む1% ギ酸溶液から上記カラムに載せ、同じ溶液中で洗浄し、90% MeOH/0.5% ギ酸で、上記ESI源へと直接溶出した。コーン電圧(cone voltage)は、60eVであり、衝突セル上の電圧は、20Vであった。800〜3000uの質量範囲にわたってスペクトルを獲得し、1サイクルあたり5秒にわたってデータを蓄積した。
【0067】
トリプシン消化のために、PRL−1−WT、PRL−1−H166AおよびPRL−1−C170S−C171Sを、20mM β−メルカプトエタノールありまたはなしの炭酸水素アンモニウム中(pH8.2)で1mg/mLに希釈し、トリプシン(Promega)を、1:50比のプロテアーゼ:タンパク質(w/w)で添加し、37℃において16時間にわたってインキュベートした。完全な消化を確認するために、種々の時点でのサンプルをSDS−PAGEで泳動し、銀染色で可視化した。MS分析と組み合わせて、キャピラリーHPLC分離を、10μL/分での勾配を発生させるクロマトグラフ(Waters capLC XL,Milford,MA)とともに、Micro−Tech Scientific(Sunnyvale,CA)によって充填されたZorbax SBC18 RPカラム(5cm×0.32mm内径,3.5μm ビーズサイズ、300Å ポアサイズ)を使用して行った。20%から80% Bへの直線勾配を、120分かけて適用した。溶媒は、以下であった:A 99% H2O、1% MeOH、B 99% MeOH、1% H2O(ともに0.08% ギ酸)。アルゴンを、上記ビームを約20%まで減衰させる圧力で、上記衝突セルに入れた。このことは、供給調節器での16psiに、または衝突セル付近にあるペニングゲージでの5.3×10−5mBarに対応する。上記衝突セルを、最大移動のために8Vで操作し、スペクトルを、250〜2000uの範囲にわたって獲得し、1サイクルあたり8秒間のデータを蓄積した。
【0068】
NMR実験を行って、上記サンプルを分析した。簡潔には、2D 1H−15N HSQCスペクトルを、パルスフィールド勾配を備えた低温の三重共鳴(triple−resonance)プローブを使用して、Bruker Avance 800 MHz NMR分光器で獲得した。水抑制を、フリップ−バック(flip−back)パルスを使用して達成した。全てのスペクトルを、37℃において得、1Hにおいて2048ポイントでの16回のスキャンで獲得した。256増分もしくは128増分を、それぞれ、WTおよびC170S−C171Sの15Nにおいて集めた。サンプルを、50mM リン酸ナトリウム、100mM NaCl(pH6.5)中に調製し、5% D2Oを含めた。WT PRL−1およびPRL−1−C170S−C171Sの濃度は、それぞれ、1.0mMおよび0.8mMであった。サンプル還元を、スペクトル獲得の前に、10mM DTTを少なくとも24時間添加することによって達成した。1H化学シフトを、D2O中の外部DSS標準に関して参照した(29)。1Hに対する間接的な参照は、13Cおよび15Nに対して決定し、このことから、比13C/1H=0.251449530および15N/1H=0.101329118を想定した。コンピュータープログラムnmrPipeおよびSparkyを、データ処理およびスペクトル分析のために使用した。
【0069】
(実施例6)
上記MAPタグモチーフが金属結合を完全に担うことを確認するために、本発明者らは、NCCを合成した。上記NCCペプチドを合成し、精製し、上記ペプチドを、金属充填IMAC樹脂とともにインキュベートした。錯体形成を、陰イオンモードで作動するエレクトロスプレーイオン化質量分析法(ESI−MS)を使用して確認した(図3)。
【0070】
上記NCCペプチドにおいて、吸収シグナルは、PRL−1中に埋まった上記ペプチド−金属錯体について観察したことを模倣するが、上記錯体は、上記トリペプチドの中により露出している。上記可視範囲における吸収は、そのpHが6から7.4へと上昇する場合に強くなる。酸性化は、そのシグナル強度を減少させ、より塩基性のpHに戻った場合に、完全なシグナルが戻ることはない。このことは、上記平面四角形の幾何的配置からいくつかの金属の不可逆的な損失が起こり得ることを示す。上記pHを10へと上昇させると、第5の配位子である、おそらく水もしくはヒドロキシドが、配位し、四角錐配置を生じることが示唆される。pH7.4において、この配位子は、おそらく高度に交換可能であり、吸収分光法によって検出するにはあまりに一過性過ぎる。上記交換可能な配位子は、上記バルク溶媒の常磁性緩和への達成方法を提供し、MRIによって検出され得る(図4)。さらに、上記金属塩を、上記ペプチドを含む水溶液へと混合することによって、上記金属を添加した。条件に依存して、上記ペプチドの吸収スペクトルは、金属移動を介して形成された上記錯体に類似であったか、または全長タンパク質について得られたものとは全く異なっていたかのいずれかであった。上記異なるスペクトルは、上記金属が、この場合、硫黄によってのみ配位されている可能性があることを明らかにする。上記NCCペプチドは、金属結合を付与するに十分であるが、生成される錯体のタイプは、上記金属が上記ペプチドに導入される方法に依存する。金属交換反応は、上記金属を、上記NCCトリペプチドに組み込んで、極めて高親和性の平面四角形の錯体を生成するための効率的な方法を提供する。
【0071】
NGHのGGH様金属錯体は、図1の構造1に示される。本発明者らは、上記金属結合NCC錯体について2つの考えられる化学的構造を想定した。これらを、図1の構造2および構造3に示す。構造2は、いくつかの脱プロトン化骨格N原子によって上記金属の配位を伴うGGH様配置に対応する。この場合、Hisに由来するイミダゾリウム窒素は、NCCにおいて、配位した錯体における3位でCys由来の硫黄と交換される。この錯体は、3N:1S配位を有し、このことは、本発明者らの吸収スペクトルとは一致しない。上記NCCによる独特の特異的配位を確認するために、NGCも合成し、金属結合における中心的Cys側鎖の重要性を確認した。上記IMAC樹脂とともにインキュベートした上記NGCペプチドは、上記金属結合したNCCもしくはPRL−1スペクトルと似た吸収スペクトルを生じない。よって、本発明者らは、GGH様配位が利用されず、上記特定の幾何的配置において金属を結合するために遙かに低い親和性を有するはずであると結論づけた。
【0072】
構造3は、上記ペプチド単独、およびより大きなタンパク質の状況にある上記ペプチドの両方について得られたデータと完全に一致している。両方についての上記吸収スペクトルは、2つに硫黄原子が、2つの窒素原子とともに上記錯体に関与していることを示し、上記窒素原子のうちの一方は脱プロトン化され、他方は、1つのプロトンが結合されている。構造3は、N末端アミン基を伴わなくてもよく、このアミン基は、上記ペプチドおよびタンパク質のスペクトルに差異を引き起こすようである。なぜならこの原子は、上記タンパク質中のペプチド結合に埋め込まれているからである。一部はこの構造に基づいて、Q、H、K、もしくはRに由来する窒素は、上記モチーフの1位におけるNの代わりに使用され得る。
【0073】
上記金属結合部位および配位の幾何的配置を研究した。PRL−1が二価金属カチオンに結合するという知見は、興味深い。なぜなら、他のPTPaseによる特異的金属配位は報告されていなかったからである。PRL−1のC末端配列は、この酵素ファミリーの中でも独特であり、GGH様モチーフ(特定の場合においては、金属に結合し得るが、この場合では、結合しない)および上記新規なMAPタグモチーフの両方をコードする。上記GGH様モチーフの幾何的配置は、平面四角形である。濃縮したPRL−1サンプルで認められた徴的なさび色は、上記GGHペプチドの色と類似する。このことは、Ni配位が、平面四角形の幾何的配置において起こることを示唆する。本発明者らの研究から、NiがPRL−1のC末端において結合されることがさらに明らかになっている。金属配位は、PRL−1中の上記GGH様モチーフによって達成されない。なぜなら、上記おそらく必須のHisの変異は、結合に対してほとんど影響を有さないからである。両方のトリペプチドが平面四角形の配位を使用して結合するという事実にも拘わらず、各々についての化学現象は異なる。
【0074】
Ni精製WT PRL−1から得られたUV−Visスペクトルは、318nm、421nmおよび526nm付近にバンドを示す。平面四角形の幾何的配置において窒素配位子を介してNiを結合したペプチドおよび有機分子については、420nm付近の特徴的なd−dバンドが、文献中で頻繁に報告されている。Maroneyらは、最初から計算を行い、平面四角形の幾何的配置を使用して、Ni(II)およびCu(II)を配位する合成ペプチド模倣物のスペクトル割り当てを行った。1つのバンドを除いて、彼らの観察および計算は、PRL−1から観察されたスペクトルと非常に近く一致している。別個に、NiSODの還元した平面四角形形態を、密度汎関数理論を使用して分析した。その結果は、類似の関連する特徴を示す。これらシステムにおける上記バンドの位置およびそれらの相対強度の傾向は、上記PRL−1タンパク質スペクトルにおいて十分に保存されている。これらおよび他の研究に基づいて、526nmバンドは、Nixy−−>Nix2−y2転移から生じるようである。同様に、上記421nmバンドは、Niz2−−>Nix2−y2転移からくる可能性がある。強い上記318nmバンドは、上記配位子−金属電荷転移バンド(LMCT)を反映する。上記スペクトルの分解から、この吸収が、306nm、325nm、および372nmにおける3つのバンドが複合しているようであることが明らかにされている。他のガイドとしての構造を使用して、306nmにおける最も鋭く最も強いバンドが、硫黄原子からの転移に対応し、他の2つのバンドは、窒素供与またはおそらくより弱い硫黄供与から生じる可能性がある。
【0075】
一連のC末端変異体の中で、C170およびC171を含む変異のみが、これら波長でのシグナルを破壊した。最も顕著なことには、上記LMCTに対応する318nmエンベロープ(envelope)は、上記システイン変異スペクトル中に不足している。全体の吸収プロフィールに関して、上記システイン変異体の低い300nm領域における上記小さな非常に広い吸収は、より低い波長における高強度のバンド、または散乱効果からの人工産物であるようである。なぜなら、それは、上記318nmの成分バンドのうちの1つであると不正確に位置づけられているからである。Ni d軌道(421nm、526nm)内の転移に特徴があり得るバンドはまた、硫黄が酸素によって置換される場合には存在しない。元のバンドは存在せず、さらなるバンドは、C170もしくはC171のいずれかの変異の際に吸収スペクトル中に現れる。また、これら残基に関わるジスルフィド結合は、上記MSデータにおいて観察されない。よって、両方のシステインが、金属配位に直接関与するようである。本発明者らのICP−MSデータは、C170およびC171に由来する両方の硫黄原子が、堅いNi結合に必要とされることをさらに示す。なぜなら、いずれの硫黄原子も、単独では十分ではないようであるからである。
【0076】
PRL−1によって生成されるスペクトル特徴はまた、Niが、残っている2つの位置における窒素原子によって配位されることを示唆する。頻繁には、Hisに由来するイミダゾリウムのNもしくは骨格アミドのNは、金属(例えば、Zn、CuおよびNi)を配位することが観察される。平面四角形の幾何的配置を使用して、Niを結合するモデルペプチドおよびタンパク質において、上記繋がる窒素基のうちのいくつかは、脱プロトン化され、負に荷電した種が、上記金属を結合する。上記526nmにおける吸収バンドは、PRL−1が、脱プロトン化窒素を介してNiを配位することを示すのに対して、上記PRL−1スペクトルにおいて生じた421nm吸収は、単独でプロトン化された窒素による配位を反映する。PRL−1−H166Aを作製するための上記GGH様モチーフにおけるHisの変異は、このヒスチジンの側鎖が、Ni結合に実質的に影響を及ぼさないことを明らかにした。PRL−1は、H166に加えて、3つの他のHisをコードする。H23、H64およびH103は、上記配列の異なる領域に位置し、変性条件下で堅く結合したままである可能性は低い。上記金属を放出するために上記タンパク質を解こうとする試みは、高温で数時間にわたる高濃度の変性剤での激しい処理を必要とした。化学的変性および熱変性は、Niのゆっくりとした放出のみを可能にし、CDデータは、Niが放出される十分前にPRL−1が完全に解かれることを示すので、上記金属は、上記タンパク質の短いセグメントにおいてクラスター形成した原子によって配位されるようである。
【0077】
上記GGHタイプペプチドは、極めて堅い結合(Kd 約10−17〜10−18)を達成する。このことは、上記モチーフ内の脱プロトン化骨格アミド窒素によって一部達成される。PRL−1について観察される極めて堅い結合および窒素原子からNi連結への推定される寄与に鑑みれば、PRL−1に由来する骨格アミドは、金属配位に関与し得る。いずれかのシステインの非存在が、結合を破壊し、短縮が両方の側鎖および骨格アミドを除去するので、金属交換反応が、変異誘発を使用して、C170およびC171に隣り合うアミド窒素に関わるという仮説を試験することは、可能でない。構造研究は、さらなる配位原子を同定する最もよい方法であり、これら研究は、本発明者らの実験室で進行中である。
【0078】
(実施例7)
生理学的pHにおけるCu−NCCを、生理食塩水バッグによって囲まれたチューブの中に入れ、University of Kansas Medical CenterにあるHoglund Brain Imaging Centerにおいて、標準的MRIを使用して画像化した(図4)。図4において、中心にある明るいシグナルは、上記Cu−NCC錯体に対応するのに対して、あまり明るくないシグナルは、上記生理食塩水溶液に対応する。黒色は、上記チューブおよび空気に対応する。
【0079】
従って、本発明は、言及した目的および利点、ならびにそこに固有のものを得るために十分に適合される。本発明は、その趣旨もしくは本質的な特徴から逸脱することなく、他の特定の形態において具現化され得る。記載される実施形態は、単なる例示として全ての観点において考慮されるべきであり、限定として考慮されるべきではない。本発明の範囲は、従って、前述の説明によってではなく、添付の特許請求の範囲によって示される。特許請求の範囲の等価物の意味および範囲内に入る全ての変更は、特許請求の範囲の範囲内に包含されるべきである。本明細書で記載される全ての参考文献は、具体的参照によって本明細書に参考として援用される。
【特許請求の範囲】
【請求項1】
明細書中に記載の発明。
【請求項1】
明細書中に記載の発明。
【図1】

【図2】

【図3】
【図4】
【図5】

【図6A】
【図6B】
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【公開番号】特開2013−107901(P2013−107901A)
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2013−37027(P2013−37027)
【出願日】平成25年2月27日(2013.2.27)
【分割の表示】特願2011−509660(P2011−509660)の分割
【原出願日】平成21年5月13日(2009.5.13)
【出願人】(508176441)ユニバーシティ・オブ・カンザス (9)
【Fターム(参考)】
【公開日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2013−37027(P2013−37027)
【出願日】平成25年2月27日(2013.2.27)
【分割の表示】特願2011−509660(P2011−509660)の分割
【原出願日】平成21年5月13日(2009.5.13)
【出願人】(508176441)ユニバーシティ・オブ・カンザス (9)
【Fターム(参考)】
[ Back to top ]