
S−トリアジンおよびジアジンを分解するための酵素および方法
本発明は、アトラジンなどのS−トリアジンおよびジアジンを分解するためのポリペプチドに関する。また、これらのポリペプチドをコードするポリヌクレオチドも提供する。本発明は、S−トリアジンおよびジアジンのバイオレメディエーションにおける、これらのポリヌクレオチドおよびポリペプチドの使用にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、アトラジンなどのS−トリアジンおよびジアジンを分解するためのポリペプチドに関する。また、これらのポリペプチドをコードするポリヌクレオチドも提供する。本発明は、S−トリアジンおよびジアジンのバイオレメディエーションにおける、これらのポリヌクレオチドおよびポリペプチドの使用にも関する。
【背景技術】
【0002】
最近の集約農業の実行は、トリアジン除草剤などの効果的な化学的ペストコントロール剤の使用によって促進される。例えば、アトラジン(6−クロロ−N2−エチル−N4−イソプロイル−1,3,5−トリアジン−2,4−ジアミン)は、非常に効果的な発芽前および後のトリアジン除草剤であり、1958年に最初に導入されてから、広葉雑草種のコントロール用に広範に使用されてきた(Tomlin、2006)。
【0003】
環境的に意味のある濃度のアトラジンは、因果関係として脊椎動物種において内分泌機能異常に関連し(例えば、アフリカツメガエルの男性性徴消失(demasculination))(Hayesら、2002、2003および2006)、アトラジンが発癌性である可能性があることが示唆されている(Huff、2002;HuffおよびSass、2007)。さらに、それらの幅広い特異性のため、アトラジンおよび関連のトリアジン除草剤は、非標的の光合成種へのそれらの毒性作用により、環境の損害を引き起こす可能性を有する。
【0004】
アトラジンは、環境中で移動性および持続性の両方である。アトラジンの環境的半減期は、4および57週間の間と見積もられており(Belluckら、1991)、アトラジンは、いくつかの国において、地表水および地下水の両方で検出されている(ThurmanおよびMeyer、1996;van der Meer、2006;Gavrilescu、2005)。
【0005】
いくつかの遺伝子/酵素系が、炭素および窒素源として、トリアジン農薬の異化を可能にする原核生物において進化している。もっとも完全に特性評価されたこれらの経路は、シュードモナス菌種ADPからもともと単離された(Mandelbaumら、1995;de Souzaら、1995)、伝播性pADP1プラスミド(Martinezら、2001)に由来するatzABCDEF遺伝子によりコードされる。アトラジンおよびシマジン(6−クロロ−N2,N4−ジエチル−1,3,5−トリアジン−2,4−ジアミン)(de Souzaら、1996)は、atzA、atzBおよびatzCによりコードされるアミドヒドロラーゼファミリー酵素によって、連続的に脱塩素化および脱アルキル化され(de Souzaら、1996;Boundy−Millsら、1997;Sadowskyら、1998)、シアヌル酸を産生し、これは、次いでatzD、atzEおよびatzFによってコードされる、経路中の残りのヒドロラーゼによって、アンモニアおよび二酸化炭素にミネラル化される(Frucheyら、2003;Chengら、2005;Shapirら、2005a)。
【0006】
AtzABCDEFアトラジン分解異化経路のAtzA酵素は、わずか25.4%同一であるにもかかわらず、TrzNトリアジン分解酵素と頻繁に置き換えられ(Sajjaphanら、2004)、これはAtzAと重複する活性を有する(Shapirら、2005b)。TrzNは、亜鉛依存性アミドヒドロラーゼファミリー酵素であり(Shapirら、2006)、トリアジン化合物から塩化物、フッ化物、S−メチル、S(O)−メチルおよびシアノ基の加水分解の置換にを担う(Shapirら、2005b)。これは、TrzNが、幅広くs−トリアジンを標的にする一方、AtzAは、ハロゲン化したs−トリアジンを解毒することのみに使用できることを意味する。TrzNは、クロロ−s−トリアジン(例えば、アトラジン、プロパジンおよびシマジン)、メチルオキシ−s−トリアジン(例えば、アトラトン、シメトンおよびプロメトン)およびメチルチオ−s−トリアジン(例えば、アメトリン、プロメトリンおよびシメトリン)除草剤を標的にする。
【0007】
TrzNはまた、AtzAの5秒−1と比較して、同程度のAtzA2.1秒−1の触媒定数を有するが、アトラジンについてははるかにより低いKmを有することが報告されている(AtzAについて100μMと比較して20μM)(Shapirら、2006)。したがって、AtzAは、3.3×104のアトラジンについてKcat/Kmを有するが、TrzNは、アトラジンについて1×105のKcat/Kmを有し、TrzNは、AtzAより触媒作用が効率的な酵素であることを実際に示す。
【0008】
しかし、AtzAと違い、TrzNは、大腸菌などの異種宿主において有意量で発現させるのは難しいことがわかっている。TrzNを分子シャペロンGroELとともに共発現させたとき、10mg.mL−1より少ない最大収率を大腸菌から得たが(Shapirら、2006)、シャペロンの非存在ではわずか560μg.mL−1の最大収率であった(Shapirら、2005b)。
【0009】
バイオレメディエーションは、潜在的に有害な残留農薬の環境への影響を改善するための新興のアプローチである(Alcaldeら、2007)。成功したバイオレメディエーション戦略の1つは、酵素的バイオレメディエーションのものであり、ここでは単離または半精製した酵素を、農薬の毒性を大幅に低下させるようなやり方で、毒性農薬を異化または修飾するために使用する(Paralesら、2002;Sutherlandら、2004)。酵素的バイオレメディエーションは、生微生物の使用よりも多くの利点を有する;GM生物またはインタクトなDNAの環境中への放出があり、使用する酵素は一般的に素早く(ほんの数時間の適用時間を必要とする)、適用後の制限された予測可能な持続性を有する(Alcaldeら、2007)。
【0010】
しかし、バイオレメディエーションに採用する酵素の要件は、いくらか厳しく、高い触媒活性、低いKm、拡散性の補助因子への非依存、および環境条件(pH、温度、塩濃度など)の範囲に対して一般的に頑強なタンパク質が必要とされる。酵素はまた、大腸菌などの典型的な発酵生物中で、高い可溶性の活性タンパク質として発現しなければならず、この点においてTrzNは不適当である。
【発明の概要】
【発明が解決しようとする課題】
【0011】
アトラジンの潜在的に大きな環境への足跡が心配されるが、農業においてその継続的な使用が所望される。したがって、環境のダメージに対し、アトラジンおよび他のs−トリアジンならびにジアジンの潜在力を除去するか低下させるためのさらなる方法が必要である。
【課題を解決するための手段】
【0012】
本発明者らは、TrzNをコードするポリヌクレオチド、または強化した特性を有するその変異体を同定した。
【0013】
第1の態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解するポリペプチドをコードする、単離された、かつ/または外因性のポリヌクレオチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリヌクレオチドを提供する。
【0014】
好ましい実施形態では、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが、可溶性の生物学的活性形態で産生される。
【0015】
さらに特に好ましい実施形態では、ポリヌクレオチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードする。別の実施形態では、ポリヌクレオチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含むポリペプチドをさらにコードする。
【0016】
好ましい実施形態では、ポリヌクレオチドは、以下のヌクレオチドの置換の1つもしくは複数、またはそこに対応するヌクレオチド位置での置換を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む;T5C、C39A、C76A、C84A、T87C、C101A、T108A、T108A、G112A、A127G、C135T、A157T、C165T、G168A、C180T、C189T、A200T、C207T、G210A、G225T、A228G、C229T、T240C、A250C、C268A、G270A、A271T、T273A、C279T、A296G、A302G、A303G、A314G、C315A、T317C、T320C、A326C、A333G、T336C、C346T、G357A、A367G、C372T、C375A、C381T、T384C、C391A、C391G、C391T、T392C、T392A、T392G、G393C、T399C、C410T、C411A、C411T、A414G、A418C、T423C、T426A、A432T、C438T、C449G、C454T、T466C、T468C、T471C、C474T、G475A、C476T、A481G、C483T、G489A、G489T、T498C、T531A、A537G、A540G、A545G、T546G、G548A、T555C、T555C、C564T、G567A、G567A、G568A、G569A、G573C、T579C、A584T、G589A、A600G、T618C、T618C、T627C、C628G、G630A、C633T、G637A、T639C、T639C、A654G、G660A、G660T、T663C、C675A、G681T、C686T、C690A、C696T、G705A、A723G、C727G、T728C、T728G、G729C、G736A、G737C、G737A、G737T、T738G、T738C、T738C、G745A、G753C、G768A、T774A、C807A、T840A、A843G、A852T、A855G、C867T、T879C、G880A、G880T、G880C、C881T、G882T、C885T、G897A、T900C、T906A、A928G、A938T、T941C、C957A、T959A、C972T、T978A、C981T、C993T、C999T、C1003A、C1003T、T1011C、G1048A、G1048T、G1048C、A1049T、A1049G、G1053A、A1059G、A1086G、G1094A、T1101C、T1101G、C1128T、A1152G、G1176T、C1186A、C1186T、T1196C、C1203T、G1221A、C1223T、C1236T、G1248T、G1270A、C1278T、T1286A、T1305C、G1309A、C1321T、A1326G、C1329T、C1329T、C1332T、C1344A、C1351AおよびG1353T。
【0017】
ある実施形態では、ポリヌクレオチドは、以下のアミノ酸の置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する、配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;
I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【0018】
さらなる実施形態では、ポリヌクレオチドは、以下のアミノ酸の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する、配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;M82、W85、L86、M92、L131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325およびS329。
【0019】
特に好ましい別の実施形態では、ポリヌクレオチドは、配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含む。
【0020】
さらに好ましい実施形態では、ポリヌクレオチドは、
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置に、アスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置に、スレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含むポリペプチドをコードする。
【0021】
別の実施形態では、ポリヌクレオチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0022】
さらに別の実施形態では、ポリヌクレオチドは、以下のヌクレオチド置換もしくは置換グループの1つ、またはそこに対応するヌクレオチド位置(複数可)での置換(複数可)を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む;
i)T468C、
ii)T468C、A938T、
iii)A200T、G210A、T468C、
iv)T468C、C476T、G753C、
v)T468C、C476T、T728C、
vi)T468C、1048T、
vii)T384C、T468C、G569A、G681T、
viii)T468C、G475A、
ix)C279T、T468C、C1223T、C1329T、
x)C76A、T468C、A481G、
xi)C39A、C101A、T468C、T639C、G737C、G1048T、
xii)C410T、A418C、T468C、A600G、
xiii)T468C、G705A、C1003A、
xiv)T468C、G573C、
xv)T468C、C474T、C628G、C1278T、
xvi)T399C、T468C、G880A、T900C、
xvii)T468C、C1236T、
xviii)T87C、A367G、T468C、
xix)T468C、G1176T、
xx)C135T、T468C、C1344A、
xxi)T468C、A852T、
xxii)T468C、T738C、
xxiii)C454T、T468C、
xxiv)A200T、T468C、G1309A、
xxv)T468C、G489T、G745A、
xxvi)C410T、T468C、
xxvii)T468C、G736A、
xxviii)G225T、C268A、A414G、T468C、T627C、C1321T、
xxix)A432T、T468C、T471C、C476T、T728C、G1053A、C1351A、
xxx)A303G、C449G、T468C、C476T、C686T、C690A、T728C、C1128T、
xxxi)A200T、G210A、C372T、T468C、A654G、C1003A、
xxxii)C180T、A200T、A302G、C375A、T399C、C411T、T468C、A540G、G880A、T900C、
xxxiii)C229T、T468C、G705A、C1003A、
xxxiv)T317C、T468C、A481G、T531A、G753C、T906A、T978A、C1003A、A1326G、
xxxv)A127G、T320C、T468C、C476T、G753C、G1048T、
xxxvi)A157T、C410T、A418C、T468C、A545G、G568A、A600G、C628G、G630A、G1048T、C1332T、
xxxvii)T468C、C476T、A723G、C1003A、G1048T、C1329T、
xxxviii)C84A、T399C、T468C、C483T、G880A、T900C、G1048A、
xxxix)T468C、T498C、C628G、C885T、A1086G、G1270A、
xl)T384C、T468C、C476T,G569A、G753C、A1152G、
xli)T336C、T468C、C476T、A537G、C564T、
xlii)A228G、T240C、T468C、C628G、G880A、G1048T、G1094A、
xliii)T273A、A367G、C381T、T399C、T468C、A481G、G880A、T900C、
xliv)A200T、C207T、G210A、T468C、C476T、G1048T、A1059G、
xlv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1236T、
xlvi)A200T、G210A、C346T、T468C、C476T、T579C、T728C、C1278T、
xlvii)C438T、T468C、C476T、A855G、
xlviii)G168A、T426A、T468C、C476T、C628G、C1203T、T1305C、
xlix)T468C、C476T、T663C、C1236T、
l)T384C、T468C、C476T、T728C、A928G、C1003A、C1186A、
li)T5C、C315A、T468C、C476T、G589A、G681T、T728C、G897A、C1003A、
lii)C279T、T468C、C476T、C1003A、
liii)A250C、A314G、T468C、C476T、
liv)A200T、G210A、T468C、T555C、G880A、T900C、
lv)T468C、C476T、T546G、C696T、C1003A、G1048T、
lvi)A200T、G210A、T468C、C476T、T728C、T1101G、
lvii)G112A、C165T、T468C、C476T、T618C、G753C、C999T、T1101C、
lviii)T423C、T468C、C476T、G489A、A584T、G753C、G1048T、
lix)T466C、T468C、C474T、C628G、G1048T、
lx)T108A、A200T、G210A、G357A、T468C、G1048T、A1059G、C1278T
lxi)T468C、C476T、G567A、G753C、G1048T、
lxii)A200T、G210A、A296G、T468C、C476T、G637A、T728C、C1003A、
lxiii)C189T、A326C、T468C、C476T、T941C、T959A、A1059G、C1186T、T1196C、G1248T、T1286A、
lxiv)T468C、C476T、C1003A、
lxv)A200T、G210A、C279T、T468C、C476T、T879C、C1003A、G1048T、
lxvi)G112A、C165T、T392C、T468C、C476T、T618C、G660A、C675A、G753C、C807A、C993T、C999T、T1101C、T1305C、
lxvii)A271T、T392C、T468C、C476T、G753C、G880A、G1048T、G1094A、C1186A、G1221A、
lxviii)A228G、T240C、T468C、G548A、C628G、G1048T、C1332T、
lxix)T108A、A200T、G210A、T423C、T468C、C476T、G567A、G1048T、
lxx)T384C、T468C、C476T、C628G、C867T、G880A、T900C、C981T、G1048A、A1326G、
lxxi)A200T、C207T、G210A、T468C、C476T、T840A、T900C、G1048A、
lxxii)T468C、C476T、C633T、G660T、A723G、C1003A、G1048T、C1329T、
lxxiii)A228G、T240C、G270A、T468C、C628G、880A、G1048T、
lxxiv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1003T、C1236T、
lxxv)A228G、T240C、T468C、C628G、G880A、C957A、C1003A、または
lxxvi)A200T、G210A、C411A、T468C、T555C、T639C、A654G、G768A、T774A、C972T、C1003A、T1011C、G1353T。
【0023】
ある実施形態では、細菌細胞中で少なくとも2回、より好ましくは少なくとも5回発現させると、ポリペプチドの量は、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも産生される。
【0024】
さらなる実施形態では、細菌細胞中で少なくとも2回、より好ましくは5回発現させると、可溶性の生物学的活性ポリペプチドの量は、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも産生される。
【0025】
代替的な実施形態では、大腸菌株BL21λDE3中で発現させ、アンピシリン200μg.mL−1、IPTG1μMを補充したLB寒天プレート上で培養し、アトラジン1mg.mL−1(90%アトラジンw/w)を浸透させるとき、コロニー付近の培地の清澄化は、培養から約8日以内、より好ましくは約6日以内、さらにより好ましくは約2日以内に検出することができる。かかる条件下のポリヌクレオチドの発現は、実施例の部でさらに詳細に記載する。
【0026】
好ましい実施形態では、ポリペプチドは、配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍、より好ましくは少なくとも5倍、さらにより好ましくは7倍のアトラジン加水分解活性を有する。
【0027】
好ましい実施形態では、ポリペプチドは、配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍、より好ましくは少なくとも5倍のシマジン加水分解活性を有する。
【0028】
本発明のポリヌクレオチドによりコードされるポリペプチドによって加水分解できるs−トリアジンの例には、これに限定されないが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンが挙げられる。
【0029】
細菌細胞は、本発明のポリヌクレオチドによってコードされるポリペプチドを産生できる任意の細胞であってよい。好ましい実施形態では、細菌細胞は、大腸菌である。適した大腸菌株の例には、これに限定されないが、BL21λDE3(ATCC寄託番号PTA−2657)、JM109およびDH10βが挙げられる。
【0030】
さらに好ましい実施形態では、ポリヌクレオチドは、細胞中でポリヌクレオチドの発現を導くことができるプロモーターに作動可能に連結されている。
【0031】
別の実施形態では、ポリヌクレオチドは、少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質をコードする。少なくとも1つの他のポリペプチドは、例えば、本発明のポリペプチドの安定性を強化するポリペプチド、細菌細胞もしくは酵母細胞などの細胞からの融合タンパク質の分泌を促すポリペプチド、または融合タンパク質の精製を補助するポリペプチドであってよい。
【0032】
別の態様では、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。
【0033】
また、本発明のポリヌクレオチド、および/または本発明のベクターを含む宿主細胞も提供する。
【0034】
ある実施形態では、宿主細胞は、シャペロンをコードする外因性ポリヌクレオチドをさらに含む。
【0035】
本発明の宿主細胞の例には、これに限定されないが、細菌細胞、酵母細胞または植物細胞が挙げられる。
【0036】
さらなる態様では、本発明は、本発明の少なくとも1つの細胞を含むトランスジェニック植物を提供する。
【0037】
さらに別の態様では、本発明は、本発明の少なくとも1つの細胞を含む、トランスジェニック非ヒト動物を提供する。
【0038】
さらなる態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解する、実質的に精製された、および/または組換えポリペプチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリペプチドを提供する。
【0039】
好ましい実施形態では、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが、可溶性の生物学的活性形態で産生させる。
【0040】
さらに特に好ましい実施形態では、ポリペプチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含む。別の実施形態では、ポリペプチドはさらに、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含む。
【0041】
本発明のポリペプチドによって加水分解できるs−トリアジンの例には、これに限定されないが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンが挙げられる。
【0042】
ある実施形態では、ポリペプチドは、少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質である。少なくとも1つの他のポリペプチドは、例えば、本発明のポリペプチドの安定性を強化するポリペプチド、細菌細胞もしくは酵母細胞などの細胞からの融合タンパク質の分泌を促すポリペプチド、または融合タンパク質の精製を補助するポリペプチドであってよい。
【0043】
別の実施形態では、ポリペプチドは、固体支持体に固定化されている。
【0044】
別の態様では、本発明は、本発明の宿主細胞の抽出物、本発明の植物および/または本発明の動物を提供し、抽出物は、本発明のポリペプチドを含む。
【0045】
さらなる態様では、本発明は、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、および/または本発明の抽出物、および1つもしくは複数の許容される担体を含む組成物を提供する。
【0046】
さらなる態様では、本発明は、s−トリアジンまたはジアジンを加水分解するための方法であって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物と、s−トリアジンまたはジアジンを接触させるステップを含む方法を提供する。
【0047】
ある実施形態では、土壌、水、生体物質またはそれらの組合せからなる群から選択される試料。
【0048】
別の態様では、本発明は、対象においてs−トリアジンまたはジアジンより引き起こされた毒性を処理する方法であって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物を対象に投与するステップを含む方法を提供する。
【0049】
別の態様では、本発明は、対象においてs−トリアジンまたはジアジンにより引き起こされた毒性の処理用の薬剤を製造するための、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物の使用を提供する。
【0050】
別の態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解できるポリペプチドを産生する方法であって、ポリペプチドをコードするポリヌクレオチドの発現を可能にする条件下で、前記ポリペプチドをコードする本発明の宿主細胞、または前記ポリペプチドをコードする本発明のベクターを培養するステップと、発現したポリペプチドを回収するステップとを含む方法を提供する。
【0051】
なおさらなる態様では、本発明は、宿主細胞を検出するための方法であって、
i)細胞または細胞集団を、細胞(複数可)によるポリヌクレオチドの取込みを可能にする条件下で、本発明のポリヌクレオチドと接触させるステップと、
ii)ステップi)からの細胞またはその子孫細胞を、s−トリアジンまたはジアジンにさらすことにより宿主細胞を選択するステップと
を含む方法を提供する。
【0052】
ある実施形態では、ポリヌクレオチドは、本発明のポリペプチドをコードする。
【0053】
ある実施形態では、ポリヌクレオチドは、本発明のポリヌクレオチドを含む第1オープンリーディングフレーム、および本発明のポリヌクレオチドを含まない第2オープンリーディングフレームを含む。
【0054】
一実施形態では、第2オープンリーディングフレームは、ポリペプチドをコードする。別の実施形態では、第2オープンリーディングフレームは、翻訳されていないポリヌクレオチドをコードする。両場合において、第2オープンリーディングフレームは、適したプロモーターに作動可能に連結されていることが好ましい。
【0055】
好ましくは、翻訳されていないポリヌクレオチドは、触媒性核酸、dsRNA分子またはアンチセンス分子をコードする。
【0056】
好ましい実施形態では、細胞は植物細胞である。
【0057】
さらなる態様では、本発明は、s−トリアジンまたはジアジンを加水分解するためのキットであって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明によるポリペプチド、本発明の抽出物、および/または本発明の組成物を含むキットを提供する。
【0058】
さらなる態様では、本発明は、本発明のポリペプチドの結晶を提供する。
【0059】
別の態様では、本発明は、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを設計する方法であって、本発明の結晶の原子座標を使用して、s−トリアジンまたはジアジンが、候補ポリペプチドと結合する能力をコンピュータによって評価するステップと、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを選択するステップとを含む方法を提供する。
【0060】
明らかになるように、本発明の一態様の好ましい特質および特徴は、本発明の他の多くの態様に適用可能である。
【0061】
本明細書を通して、用語「含む(comprise)」、または「含む(comprises)」または「含んでいる(comprising)」などの変形は、述べられた要素、整数もしくはステップ、または要素、整数もしくはステップのグループを含有することを意味するが、他の任意の要素、整数もしくはステップ、または要素、整数もしくはステップのグループを排除しないことを理解されよう。
【0062】
本発明を、以下の非限定的実施例によって、かつ添付の図面を参照して以下に説明する。
【図面の簡単な説明】
【0063】
【図1】本発明の酵素によって分解される様々なs−トリアジンの構造を示す図である。
【図2】pETcc2::egfpの概略図である。
【図3】野生型TrzN、TrzN cc3.2および中間突然変異型の部分的な精製を示す図である。野生型TrzNに関する値は、5リットルから2.8mgを産生するShapirら(2005)から取る。分子量マーカーを含んだが(レーンM)、それらの分子量(kDa)を示す。TrzNおよびその変異体の位置は、矢印で示す。TrzNおよびその変異体の同一の精製から得た同等量(5μl)を、各レーンに添加した。
【図4】TrzN構造の概略図および絵である。TrzNは、99.6kDaのホモ二量体を形成する。各単量体は、2つのドメイン:βサンドイッチドメインおよびβ/α8バレルドメインに分割される。β/α8バレルは、上側の面上にいくつかのループの挿入を有し、これは、活性部位の入口を修飾するのに役立ち、またはさらに二量体の界面を形成する。
【図5】TrzNcc3.2の基質結合ポケットを構成する残基を示す概略図である。アトラジン、Zn2+中心およびヒドロキシルイオン(OH−)を中央に示す。基質結合ポケットを含むアミノ酸の同一性および配列位置を示す。
【図6】Zn2+を有するApo−TrzNcc3.2の飽和を示すグラフである。最大半量の活性は、ZnCl22.6μMで回復した。
【図7】活性部位のアミノ酸残基およびアミノ酸残基に配位する金属の概略図である。アトラジン、Zn2+およびヒドロキシルイオン(OH−)を中央に示し、関連残基の同一性および配列位置を示す。
【図8】提案されたTrzNの反応機構を説明する概略図である。
【図9】TrzNとアメトリンの速度定数(kcat)、および反応緩衝液のpHの間の関係を示すグラフである。
【図10】TrzNcc3.2による実地試験の結果を示すグラフである。TrzNcc3.2を添加して10.5時間後の1.5MLのホールディングダム(holding dam)中のアトラジンの枯渇。QHFSS(四角)およびCSIRO Entomology(丸)によって解析した試料。
【発明を実施するための形態】
【0064】
一般的な技法および定義
他に特に定義されない限り、本明細書で使用されるすべての技術的および科学的用語は、当業者(例えば、細胞培養、バイオレメディエーション、分子遺伝学、免疫学、免疫組織化学、タンパク質化学、および生化学における)によって一般に理解されるのと同じ意味を有すると解釈されるものとする。
【0065】
他に指定されない限り、本発明において利用される組換えタンパク質、細胞培養、および免疫学的技法は、当業者に周知の標準的な手順である。そのような技法は、出典における文献、例えば、J.Perbal、A Practical Guide to Molecular Cloning、John Wiley and Sons(1984);J.Sambrookら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbour Laboratory Press(1989);T.A.Brown(編)、Essential Molecular Biology:A Practical Approach、1巻および2巻、IRL Press(1991);D.M.GloverおよびB.D.Hames(編)、DNA Cloning:A Practical Approach、1〜4巻、IRL Press(1995および1996);およびF.M.Ausubelら(編)、Current Protocols in Molecular Biology、Greene Pub.Associates and Wiley−Interscience(1988、現在までのすべての最新版を含む);Ed HarlowおよびDavid Lane(編)Antibodies:A Laboratory Manual、Cold Spring Harbour Laboratory、(1988);ならびにJ.E.Coliganら(編)、Current Protocols in Immunology、John Wiley&Sons(現在までのすべての最新版を含む)などにわたって記載および説明されている。
【0066】
本明細書で使用するとき、用語「加水分解する」、「加水分解している」、「加水分解活性」およびその変形は、化学結合の加水分解を触媒する本発明のポリペプチドの能力をいう。好ましい実施形態では、本発明の酵素は、以下のものの1つまたは複数;デハロゲナーゼ(例えば、クロロヒドロラーゼ、フルロヒドロラーゼ、ハロヒドロラーゼ、加水分解性デコルリナーゼ(decholrinase)、加水分解性デフルロリナーゼおよび/または加水分解性デハロゲナーゼ)、メトキシヒドロラーゼおよびメチルチオヒドロラーゼである。好ましい実施形態では、酵素活性の産物が、s−トリアジンまたはジアジン基質と比べて、例えば、哺乳動物および/または魚類への毒性が弱くなり、かつ/または安定性が低くなるように、ポリペプチドは、s−トリアジンまたはジアジンを「分解する」。
【0067】
本明細書で使用するとき、用語「より高いs−トリアジンおよび/またはジアジン加水分解活性」とは、s−トリアジンまたはジアジンに関して、より高い特異的活性、触媒定数(kcat)、基質特異性(Km)および/または二次速度定数(kcat/Km)、または配列番号1として提供されるアミノ酸配列を含むポリペプチドよりも高い安定性を有する本発明のポリペプチドをいう。特異的活性は、実施例の概説のように決定することができる。
【0068】
本明細書で使用するとき、表現「そこに対応するアミノ酸位置に」および「アミノ酸番号に対応する位置に」とは、周囲のアミノ酸と比較して、相対的なアミノ酸位置をいう。例えば、いくつかの実施形態では、本発明のポリペプチドは、例えば、配列番号1に対して整列させたとき、アミノ酸の相対的な位置付けを変える、欠失または置換の突然変異を有し得る。ある実施形態では、ポリペプチドは、指定された残基番号に規定されたアミノ酸を含む。
【0069】
同様に、表現「そこに対応するヌクレオチド位置に」および「ヌクレオチド番号に対応する位置に」とは、周囲のヌクレオチドと比較して、相対的なヌクレオチドの位置をいう。例えば、いくつかの実施形態では、本発明のポリヌクレオチドは、例えば、配列番号2または配列番号4に対して整列させたとき、ヌクレオチドの相対的な位置付けを変える、欠失または置換の突然変異を有し得る。ある実施形態では、ポリヌクレオチドは、指定されたヌクレオチド番号に規定されたヌクレオチドを含む。
【0070】
本明細書で使用するとき、用語「処理している」、「処理する」または「処理」は、s−トリアジンまたはジアジンによって引き起こされる毒性の少なくとも1つの症状を和らげるまたは排除するのに十分な、処理効果のある量の本発明のポリペプチド、またはそれをコードするポリヌクレオチドを投与することを含む。
【0071】
用語「生体物質」は、本明細書において、生物起源の任意の産物を含むもっとも広範な意味で使用される。かかる産物には、これに限定されないが、ヒトのための食料産物および動物の餌が挙げられる。この産物には、水およびミルクなどの液体食材、ならびにヨーグルトなどの半流動性の食材を含む液体媒体が挙げられる。本発明はまた、固体食材、特に動物の餌にまで及ぶ。ある実施形態では、生体物質は、これに限定されないが、サトウキビ、キャノーラ種子、コムギ種子、オオムギ種子、ソルガム種子、イネ、トウモロコシ、パイナップルまたはワタ種子などの植物物質であることが好ましい。
【0072】
本明細書で使用するとき、用語「抽出物」とは、本発明の宿主細胞、植物またはトランスジェニック非ヒト動物の任意の部分をいう。この部分は、植物種子などの実体全部であってよく、または少なくとも部分的な均質化および/または精製によって得ることができる。この用語は、宿主細胞から分泌される部分を含み、したがって培養上澄み液を包含する。
【0073】
本明細書で使用するとき、用語「シャペロン」とは、細胞からのタンパク質の排出を変えるために、他のタンパク質が適切なフォールディングまたはアンフォールディングを成し遂げるのを補助する機能を有するタンパク質をいう。シャペロンは当技術分野でよく知られており、これに限定されないが、トリガー因子(TF)などのリボソーム結合性タンパク質;Hsp70、DnaK、Hsp40、DnaJ、GrpEなどのシャペロンのHsp7Oファミリー、およびGroEL、GroES、Hsp60、Hsp10などのシャペロンのシャペロニンファミリーが挙げられる。非限定的な例として、大腸菌由来のシャペロンGroELは、シャペロンの熱ショックタンパク質60(Hsp60)クラスのメンバーであり、大腸菌のGroEオペロン由来のGroESと一緒に発現する。GroELは、凝集しやすいポリペプチド中間体の濃度、および経路から外れる凝集率を減少させるアンフォールドのタンパク質に結合することによって、タンパク質のフォールディング反応を補助し、これによって未変性の立体構造に分割するのを助ける。コシャペロニンGroEL、ならびにATP、K+およびMg2+などの補助因子は、GroEL媒介型ポリペプチドのフォールディング反応の収率をさらに増加させることが知られている。したがって、特定の実施形態では、当業者は、シャペロン媒介型(または補助型)タンパク質フォールディングを改良または強化するために、当技術分野で既知の成分、補助因子、追加のシャペロンタンパク質を含めよう。
GroELをコードするのに使用できるベクターの例には、これに限定されないが、dnaK−dnaJ−grpE−groES−groELを含むpG−KJE8、groES−groELを含むpGro7、およびgroES−groEL−tigを含むpG−Tf2が挙げられる(Nishiharaら、1998および2000)。
【0074】
s−トリアジンおよびジアジン
本明細書で使用するとき、「s−トリアジン」は、3つの炭素原子および3つの窒素原子からなる6員環複素環式芳香族環を有する化学構造を含む有機化合物である。トリアジン環中の原子は、ベンゼン環における原子と類似している。本発明の酵素によって加水分解される(分解される)s−トリアジンのタイプの例には、これに限定されないが、クロロ−s−トリアジン、フルオロ−s−トリアジン、メチルチオ−s−トリアジンおよびメチルオキシ−s−トリアジンが挙げられる。本発明の酵素によって加水分解される(分解される)いくつかのs−トリアジンの化学構造は、図1に示される。好ましい実施形態では、s−トリアジンは、次の構造を有する;
2−R1−4−R2−6−R3−1,3,5−トリアジン、式中R1が、Cl、Fl、OCH3、SCH3、S(O)CH3またはN3であってよく、R2またはR3が、OCH3、NHCH2CH2OH、NH(CH2)2CH3、NH(CH2)3CH3、NHCH2CH(CH3)2、NHCH(CH3)CH2CH3、NHC(CH3)2CN、NHC(CH3)3、NH2またはOHであってよい。
【0075】
塩素化(クロロ)s−トリアジンは、少なくとも1つの塩化物を含む。塩素化s−トリアジンの例には、これに限定されないが、アトラジン(6−クロロ−N−エチル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)(図1参照)、クロラジン(6−クロロ−N,N,N’,N’−テトラエチル−1,3,5−トリアジン−2,4−ジアミン)、シアナジン(2−[[4−クロロ−6−(エチルアミノ)−1,3,5−トリアジン−2−イル]アミノ]−2−メチルプロパンニトリル)、シプラジン(6−クロロ−N−シクロプロピル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、エグリナジン(N−[4−クロロ−6−(エチルアミノ)−1,3,5−トリアジン−2−イル]グリシン)、イパジン(6−クロロ−N,N−ジエチル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、メソプラジン(6−クロロ−N−(3−メトキシプロピル)−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、プロシアジン(2−[[4−クロロ−6−(シクロプロピルアミノ)−1,3,5−トリアジン−2−イル]アミノ]−2−メチルプロパンニトリル)、プログリナジンN−[4−クロロ−6−[(1−メチルエチル)アミノ]−1,3,5−トリアジン−2−イル]グリシン)、プロパジン(6−クロロ−N,N’−ビス(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、セブチルアジン(6−クロロ−N−エチル−N’−(1−メチルプロピル)−1,3,5−トリアジン−2,4−ジアミン、シマジン(6−クロロ−N,N’−ジエチル−1,3,5−トリアジン−2,4−ジアミン)、テルブチルアジン(6−クロロ−N−(1,1−ジメチルエチル)−N’−エチル−1,3,5−トリアジン−2,4−ジアミン)およびトリエタジン(6−クロロ−N,N,N’−トリエチル−1,3,5−トリアジン−2,4−ジアミン)、ならびに(デスエチル(desethyl)アトラジンおよびデスイソプロピル(desisopropyl)アトラジンなどのアトラジン脱アルキル化を含む)クロロ−s−トリアジン脱アルキル化の産物が挙げられる。
【0076】
メチルチオ−s−トリアジンは、少なくとも1つのチオール基を含む。メチルチオ−s−トリアジンの例には、これに限定されないが、アメトリン(N2−エチル−N4−イソプロピル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)、プロメトリン(N2,N4−ジイソプロピル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)およびシメトリン(N2,N4−ジエチル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)が挙げられる。
【0077】
メトキシ−s−トリアジンは、少なくとも1つのメトキシ基を含む。メトキシ−s−トリアジンの例には、これに限定されないが、アトラトン(N2−エチル−N4−イソプロピル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)、シメトン(N2,N4−ジエチル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)およびプロメトン(N2,N4−ジイソプロピル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)が挙げられる。
【0078】
フルオロ−s−トリアジンは、少なくとも1つのフッ素を含む。フルオロ−s−トリアジンの例は、フルオルアトラジン(2−フルオロ−4−N−エチルアミノ−6−N−イソプロピルアミノ−1,3,5−トリアジン)である。
【0079】
ある実施形態では、s−トリアジンは、少なくとも1つのN−エチル、N−イソプロピル、N−ジエチルおよび/またはN−シアノジメチルメチルアルキル側鎖を有する。
【0080】
好ましい実施形態では、ジアジンは、次の構造を有する;−2−R1−4−R2−6−R3−1,3−ピリミジンであり、式中R1が、Cl、Fl、OCH3、SCH3、S(O)CH3またはN3であってよく、R2またはR3が、OCH3、NHCH2CH2OH、NH(CH2)2CH3、NH(CH2)3CH3、NHCH2CH(CH3)2、NHCH(CH3)CH2CH3、NHC(CH3)2CN、NHC(CH3)3、NH2またはOHであってよい。ジアジンのタイプの例には、これに限定されないが、クロロ−s−ジアジンおよびフルオロ−s−ジアジンが挙げられる。除草剤であるジアジンの例は、ブロマシル(5−ブロモ−3−sec−ブチル−6−メチルウラシル)である。
【0081】
ポリペプチド
「実質的に精製された」または「精製された」とは、1つまたは複数の脂質、核酸、他のポリペプチド、またはポリペプチドがその天然状態で伴っている他の混入分子から分離されたポリペプチドを意味する。実質的に精製されたポリペプチドは、ポリペプチドが天然に伴っている他の成分を、少なくとも60%、より好ましくは少なくとも75%、より好ましくは少なくとも90%含まないことが好適である。しかし現在、本発明のポリペプチドが自然に存在する証拠はない。
【0082】
ポリペプチドとの関連において用語「組換え体」は、ポリペプチドの天然状態と比較して変化した量、または変化した速度で細胞によって、または無細胞発現系において産生されるときのポリペプチドを指す。一実施形態では、細胞は、ポリペプチドを天然に産生しない細胞である。しかし、細胞は、産生されるポリペプチドの量の変化、好ましくは増加を引き起こす非内因性遺伝子を含む細胞であり得る。本発明の組換えポリペプチドには、これが産生される形質転換(組換え)細胞または無細胞発現系の他の成分から分離されていないポリペプチド、および少なくともいくつかの他の成分から引き続いて精製される、そのような細胞または無細胞系で産生されるポリペプチドが含まれる。
【0083】
用語「ポリペプチド」および「タンパク質」は一般に、互換的に使用され、非アミノ酸基の付加によって修飾されていても、されていなくてもよい1本のポリペプチド鎖を指す。そのようなポリペプチド鎖は、他のポリペプチドもしくはタンパク質、または補助因子などの他の分子と付随することができることが理解される。用語「タンパク質」および「ポリペプチド」は、本明細書で使用する場合、本明細書に記載されるポリペプチドの変異体、突然変異体、生物学的活性断片、修飾体、類似体および/または誘導体も含む。
【0084】
ポリペプチドの%同一性は、ギャップ生成ペナルティー=5、およびギャップ伸長ペナルティー=0.3を用いてGAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)によって決定される。クエリー配列は、長さが少なくとも25アミノ酸であり、ギャップ分析は、少なくとも25アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも50アミノ酸であり、ギャップ分析は、少なくとも50アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも100アミノ酸であり、ギャップ分析は、少なくとも100アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、クエリー配列は、長さが少なくとも250アミノ酸であり、ギャップ分析は、少なくとも250アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、クエリー配列は、長さが少なくとも400アミノ酸であり、ギャップ分析は、少なくとも400アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、ギャップ分析は、それらの全長にわたって2つの配列を整列させる。
【0085】
本明細書で使用する場合、「生物学的活性断片」は、全長ポリペプチドの規定された活性を維持する、本明細書に記載されるようなポリペプチドの部分である。生物学的活性断片は、これらが規定された活性を維持する限り任意のサイズとすることができる。好ましくは、生物学的活性断片は、長さが少なくとも100、より好ましくは少なくとも400のアミノ酸である。
【0086】
好ましい実施形態は、「可溶性の生物学的活性形態」で産生されるポリペプチドに関する。当業者が理解するように、これは、細胞中で発現するときに不溶性、したがって、不活性の形態では存在しないポリペプチドをいう。さらに、生物学的活性とは、s−トリアジンおよび/またはジアジンを加水分解する能力を意味する。
【0087】
定義されたポリペプチドに関して、上記に提供したものより高い%同一性の数値は、好適な実施形態を包含することが理解されよう。したがって適用可能な場合、最低%同一性の数値を踏まえると、ポリペプチドは、該当する指定された配列番号と少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一であるアミノ酸配列を含むことが好適である。
【0088】
本明細書に記載されるポリペプチドのアミノ酸配列突然変異体は、本明細書に定義される核酸中に適当なヌクレオチドの変化を導入することによって、または所望のポリペプチドのインビトロ合成によって調製することができる。そのような突然変異体は、例えば、アミノ酸配列内の残基の欠失、挿入、または置換を含む。最終的なコンストラクトに到達するために、欠失、挿入および置換の組合せを行うことができ、ただし最終的なポリペプチド生成物は、所望の特徴を有するという条件がある。
【0089】
突然変異体(変化した)ポリペプチドは、当技術分野で既知の任意の技法を使用して調製することができる。例えば、本明細書に記載されるポリヌクレオチドは、インビトロ突然変異生成に付すことができる。そのようなインビトロ突然変異生成技法は、適当なベクターにポリヌクレオチドをサブクローニングするステップ、大腸菌(E. coli)XL−1赤色(Stratagene)などの「ミューテーター」菌株にベクターを形質転換するステップ、および適当な数を生成するために形質転換細菌を増殖させるステップを含むことができる。別の例では、本発明のポリヌクレオチドは、Harayama(1998)によって広く記載されたDNAシャッフリング技法に付される。突然変異/変化したDNAに由来する生成物は、これらが、活性の増強および/または基質特異性の変化などの所望の表現型を付与することができるかどうかを判定するために、本明細書に記載される技法を使用して容易にスクリーニングすることができる。
【0090】
アミノ酸配列突然変異体の設計において、突然変異部位の位置および突然変異の性質は、改変される特性(複数可)に依存する。突然変異の部位は、例えば、(1)実現される結果に応じて、最初に保存的なアミノ酸選択物で、次いでより急進的な選択物で置換し、(2)標的残基を欠失させ、または(3)位置づけられた部位に隣接して他の残基を挿入することによって個々に、または連続的に改変することができる。
【0091】
アミノ酸配列の欠失は一般に、約1〜15個の残基、より好ましくは約1〜10個の残基、および典型的には約1〜5個の連続した残基の範囲である。
【0092】
置換突然変異体は、取り出されるポリペプチド分子中の少なくとも1つのアミノ酸残基、およびその場所に挿入される異なる残基を有する。置換突然変異生成にとってもっとも対象となる部位は、機能に対して重要であると同定された部位を含む。対象とする他の部位は、様々な菌株または種から得られる特定の残基が同一であるものである。これらの位置は、生物活性とって重要となり得る。これらの部位、特に、3つの他のまったく同様に保存された部位の配列内に入るものは、相対的に保存された様式で置換されることが好ましい。そのような同類置換を表1に示す。
【0093】
好ましい実施形態では、突然変異体/変異体ポリペプチドは、本明細書で具体的に定義されたポリペプチドと比較したとき、1または2または3または4つの保存アミノ酸変化を有する。保存アミノ酸変化の詳細は、表1に示される。
【0094】
好ましい実施形態では、ポリペプチドは、以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【0095】
【表1】

【0096】
さらなる実施形態では、ポリペプチドは、以下のアミノ酸の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む;M82、W85、L86、M92、L131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325およびS329。これらのアミノ酸の1つまたは複数は、ポリペプチドの特異的活性、触媒定数(kcat)、基質特異性(Km)、安定性および/または二次速度定数(kcat/Km)を変えるように変化させることができる。
【0097】
さらに好ましい実施形態では、ポリペプチドは、
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンまたはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンまたはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンまたはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンまたはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含む。
【0098】
ある実施形態では、ポリペプチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含む;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0099】
より好ましくは、ポリペプチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換を有する配列番号1に提供されるアミノ酸配列を含む;
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0100】
特に好ましい実施形態では、ポリペプチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含み、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが、可溶性の生物学的活性形態で産生される。その上、この実施形態では、ポリペプチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンをさらに含むことが好ましい。
【0101】
好ましくは、他に規定されなければ、所与のアミノ酸位置で、ポリペプチドは、配列番号1として提供されるポリペプチドの対応する位置で見られるようなアミノ酸を含む。
【0102】
指定された部位のアミノ酸が、表1に提供されたアミノ酸置換と矛盾するとき、指定されたアミノ酸が好ましい。
【0103】
好ましい実施形態では、ポリペプチドは、本発明の2つの別々のポリペプチド鎖の二量体である。ポリペプチドは、ホモ二量体またはヘテロ二量体であってよい。ヘテロ二量体に関しては、2つのポリペプチド鎖は、少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも97%、より好ましくは少なくとも99%同一であることが好ましい。
【0104】
さらに好ましい実施形態では、ポリペプチドは、Zn2+またはCo2+と結合している。
【0105】
その上、さらに突然変異体を設計するとき、当業者は、TrzNcc3.2の構造に関する実施例2に提供される情報を使用することができる。
【0106】
さらに、必要に応じて、非天然アミノ酸または化学的アミノ酸類似体を、本明細書に記載されるポリペプチド中への置換または付加として導入することができる。そのようなアミノ酸として、一般に、それだけに限らないが、一般的なアミノ酸のD異性体、2,4−ジアミノ酪酸、α−アミノイソ酪酸、4−アミノ酪酸、2−アミノ酪酸、6−アミノヘキサン酸、2−アミノイソ酪酸、3−アミノプロピオン酸、オルニチン、ノルロイシン、ノルバリン、ヒドロキシプロリン、サルコシン、シトルリン、ホモシトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、フルオロ−アミノ酸、デザイナーアミノ酸、例えば、β−メチルアミノ酸、Cα−メチルアミノ酸、Nα−メチルアミノ酸など、およびアミノ酸類似体が挙げられる。
【0107】
やはり本発明の範囲内に含まれるのは、例えば、ビオチン化、ベンジル化、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護基/ブロッキング基による誘導体化、タンパク質分解切断、抗体分子または他の細胞リガンドへの連結などによって、合成中、または合成後に差次的に改変される本発明のポリペプチドである。これらの改変は、ポリペプチドの安定性および/または生物活性を増大させるのに機能を果たすことができる。
【0108】
本明細書に記載されるポリペプチドは、天然ポリペプチドの生成および回収、組換えポリペプチドの生成および回収、ならびにポリペプチドの化学合成を含めた様々な方法で生成することができる。一実施形態では、本発明の単離ポリペプチドは、ポリペプチドを生成するのに有効な条件下で、ポリペプチドを発現することができる細胞を培養し、ポリペプチドを回収することによって生成される。培養するのに好適な細胞は、本発明の組換え細胞である。有効な培養条件は、それだけに限らないが、ポリペプチド生成を可能にする有効培地、バイオリアクター、温度、pH、および酸素条件を含む。有効培地は、本発明のポリペプチドを生成するために細胞が培養される任意の培地を指す。そのような培地は一般に、同化できる炭素、窒素、およびリン酸源を有する水媒質、ならびに適切な塩、ミネラル、金属、およびビタミンなどの他の栄養分を含む。本発明の細胞は、従来の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿、およびペトリ皿において培養することができる。培養は、組換え細胞にとって適切な温度、pH、および酸素含量で実施することができる。そのような培養条件は、当業者の専門知識内である。
【0109】
ある実施形態では、本発明のポリペプチドは、細胞からのポリペプチドの分泌を導くことができるシグナル配列を含む。非常に多くのかかるシグナル配列を単離しており、これはNおよびC末端シグナル配列を含む。原核生物および真核生物のN末端シグナル配列は類似しており、真核生物のN末端シグナル配列は、細菌において分泌配列として機能できることが示されている。かかるN末端シグナル配列の例は、よく研究されている配列である、細菌のβ−ラクタマーゼシグナル配列であり、ポリペプチドの外部環境への分泌を促進するために幅広く使用されている。C末端シグナル配列の例は、大腸菌のヘモリジンA(hlyA)シグナル配列である。シグナル配列のさらなる例には、これに限定しないが、アエロリジン、アルカリホスファターゼ遺伝子(phoA)、キチナーゼ、エンドキチナーゼ、α−ヘモリジン、MIpB、プルラナーゼ、YopsおよびTATシグナルペプチドが挙げられる。
【0110】
ポリヌクレオチド
一本鎖または二本鎖の、センスもしくはアンチセンス配向または両方の組合せ、dsRNAまたはその他でのDNA、RNA、またはこれらの組合せを含めた「単離ポリヌクレオチド」とは、これが、その天然状態で付随または連結されているポリヌクレオチド配列から少なくとも部分的に分離されたポリヌクレオチドを意味する。好ましくは、単離ポリヌクレオチドは、これが天然に付随している他の成分を、少なくとも60%、好ましくは少なくとも75%、もっとも好ましくは少なくとも90%含まない。さらに、用語「ポリヌクレオチド」は、用語「核酸」と本明細書で互換的に使用される。
【0111】
ポリヌクレオチドとの関連で、用語「外因性の」は、ポリヌクレオチドの天然状態と比較して変化した量で、細胞内または無細胞発現系内に存在するときのポリヌクレオチドを指す。一実施形態では、細胞は、ポリヌクレオチドを天然に含まない細胞である。しかし、細胞は、コードされるポリペプチドの変化した、好ましくは増加した産生量をもたらしている非内因性ポリヌクレオチドを含む細胞であり得る。本発明の外因性ポリヌクレオチドは、これが存在する形質転換(組換え)細胞または無細胞発現系の他の成分から分離されていないポリヌクレオチド、および少なくともいくつかの他の成分から引き続いて精製される、そのような細胞または無細胞系で産生されるポリヌクレオチドが含まれる。
【0112】
ポリヌクレオチドの%同一性は、ギャップ生成ペナルティー=5、およびギャップ伸長ペナルティー=0.3を用いてGAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)によって決定される。他に述べられていない限り、クエリー配列は、長さが少なくとも45ヌクレオチドであり、ギャップ分析は、少なくとも45ヌクレオチドの領域にわたって2つの配列を整列させる。好ましくは、クエリー配列は、長さが少なくとも150ヌクレオチドであり、ギャップ分析は、少なくとも150ヌクレオチドの領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも300ヌクレオチドであり、ギャップ分析は、少なくとも300ヌクレオチドの領域にわたって2つの配列を整列させる。さらにより好ましくは、ギャップ分析は、2つの配列の全長にわたって2つの配列を整列させる。
【0113】
定義されたポリヌクレオチドに関して、上記に提供したものより高い%同一性の数値は、好適な実施形態を包含することが理解されよう。したがって適用可能な場合、最低%同一性の数値を踏まえると、本発明のポリヌクレオチドは、該当する指定された配列番号と少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%で、少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一である配列を含むことが好適である。
【0114】
本発明はまた、ストリンジェントな条件下で、配列番号2および/または配列番号4をコードするポリヌクレオチドにハイブリダイズするポリヌクレオチドであって、本発明のポリペプチドをコードし、および/または本明細書で規定されるような置換(複数可)を含むポリヌクレオチドにも関する。用語「ストリンジェントなハイブリダイゼーションの条件」または「ストリンジェントな条件」などは、本明細書で使用するとき、ポリヌクレオチドまたはオリゴヌクレオチドの長さとともに様々なハイブリダイゼーションの温度を含む、当技術分野で精通しているパラメータをいう。核酸ハイブリダイゼーションのパラメータは、かかる方法を集めたSambrookら(上記)およびAusubelら(上記)の文献で見つけることができる。例えば、ストリンジェントなハイブリダイゼーションの条件とは、本明細書で使用するとき、ハイブリダイゼーション緩衝液(3.5xSSC、0.02%フィコール、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mMのNaH2PO4(pH7)、0.5%SDS、2mMのEDTA)中で65℃でのハイブリダイゼーション、および各洗浄ステップは約30分で、65℃で0.2xSSC、0.1%SDS中で2回洗浄することをいう場合がある。
【0115】
特に好ましい実施形態では、ポリヌクレオチドは、配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含み、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが産生される。
【0116】
さらに好ましい実施形態では、ポリヌクレオチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードし、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが、可溶性の生物学的活性形態で産生される。その上、この実施形態では、ポリヌクレオチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含むポリペプチドをさらにコードすることが好ましい。
【0117】
本発明のポリヌクレオチドは、ここで提供される分子と比較したとき、1つまたは複数の突然変異を有することができ、この突然変異は、ヌクレオチド残基の欠失、挿入、または置換である。突然変異体は、天然に存在しても(すなわち、天然源から単離された)、合成であってもよい(例えば、核酸への部位特異的な突然変異生成を実施することによる)。
【0118】
通常、ポリヌクレオチドの単量体は、ホスホジエステル結合またはその類似体によって連結される。ホスホジエステル結合の類似体として、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホラニリデート、およびホスホラミデートが挙げられる。
【0119】
組換えベクター
本発明の一実施形態には、組換えベクターが含まれ、これは、宿主細胞内にポリヌクレオチド分子を送達することができる任意のベクター中に挿入された、少なくとも1つの本発明の単離/外因性ポリヌクレオチドを含む。そのようなベクターは、異種のポリヌクレオチド配列、すなわち、本発明のポリヌクレオチド分子に隣接して天然に見出されず、好ましくはポリヌクレオチド分子(複数可)が由来した種以外の種に由来するポリヌクレオチド配列を含有する。ベクターは、RNAであってもDNAであってもよく、原核生物であっても真核生物であってもよく、一般にトランスポゾン(US5,792,294に記載されているような)、ウイルス、またはプラスミドである。
【0120】
ある型の組換えベクターは、発現ベクターに作動可能に連結したポリヌクレオチド(複数可)を含む。作動可能に連結された、という語句は、分子が、宿主細胞内に形質転換されたとき発現され得るような様式での、発現ベクター内へのポリヌクレオチド分子の挿入を指す。本明細書で使用する場合、発現ベクターは、宿主細胞を形質転換し、指定されたポリヌクレオチド分子の発現を生じさせることができるDNAまたはRNAベクターである。発現ベクターは、宿主細胞内で複製することもできることが好ましい。発現ベクターは、原核生物であっても真核生物であってもよく、一般にウイルスまたはプラスミドである。発現ベクターには、細菌、真菌、内部寄生虫、節足動物、動物、および植物の細胞を含めた組換え細胞内で機能する(すなわち、直接遺伝子発現)任意のベクターが含まれる。本発明のベクターは、無細胞発現系においてポリペプチドを産生させるために使用することもでき、そのような系は当技術分野で周知である。
【0121】
「作動可能に連結された」は、本明細書で使用する場合、2つ以上の核酸(例えば、DNA)セグメントの間の機能的関係を指す。一般にこれは、転写される配列に対する転写調節エレメントの機能的関係を指す。例えば、プロモーターは、これが、適切な宿主細胞および/または無細胞発現系において、本明細書に定義されるポリヌクレオチドなどのコード配列の転写を刺激または調節する場合、このコード配列に作動可能に連結されている。一般に、転写される配列に作動可能に連結されているプロモーターの転写調節エレメントは、転写される配列に物理的に連続している、すなわち、これらはシス作用である。しかし、エンハンサーなどのいくつかの転写調節エレメントは、これらが転写を増強するコード配列に物理的に連続している、またはごく接近して位置している必要はない。
【0122】
特に、本発明の発現ベクターは、組換え細胞と適合性であり、本発明のポリヌクレオチド分子の発現を制御する制御配列、例えば、転写制御配列、翻訳制御配列、複製起点、および他の制御配列などを含有する。特に、本発明の組換え分子は、転写制御配列を含む。転写制御配列は、転写の開始、伸長、および終止を制御する配列である。特に重要な転写制御配列は、転写開始を制御するもの、例えば、プロモーター、エンハンサー、オペレーター、および抑制因子配列などである。適当な転写制御配列には、本発明の組換え細胞のうちの少なくとも1つにおいて機能することができる任意の転写制御配列が含まれる。様々なそのような転写制御配列は、当業者に既知である。好適な転写制御配列として、細菌、酵母、節足動物、線虫、植物、または動物細胞において機能するもの、例えば、それだけに限らないが、tac、lac、trp、trc、oxy−pro、omp/lpp、rrnB、バクテリオファージλ、バクテリオファージT7、T71ac、バクテリオファージT3、バクテリオファージSP6、バクテリオファージSP01、メタロチオネイン、α−接合因子、ピキア(Pichia)アルコールオキシダーゼ、アルファウイルスサブゲノムプロモーター(シンドビスウイルスサブゲノムプロモーターなど)、抗生物質耐性遺伝子、バキュロウイルス、ヘリオチス・ゼア(Heliothis zea)昆虫ウイルス、ワクシニアウイルス、ヘルペスウイルス、アライグマポックスウイルス、他のポックスウイルス、アデノウイルス、サイトメガロウイルス(中間初期プロモーターなど)、シミアンウイルス40、レトロウイルス、アクチン、レトロウイルスロングターミナルリピート、ラウス肉腫ウイルス、熱ショック、リン酸および硝酸転写制御配列、ならびに原核生物細胞または真核細胞において遺伝子発現を制御することができる他の配列などが挙げられる。
【0123】
宿主細胞
本発明の別の実施形態は、本明細書で記載の1つもしくは複数の組換え分子またはその子孫細胞より形質転換させる宿主細胞を含む。細胞内へのポリヌクレオチド分子の形質転換は、細胞内にポリヌクレオチド分子を挿入することができる任意の方法によって実現することができる。形質転換技法には、それだけに限らないが、トランスフェクション、電気穿孔、微量注入、リポフェクション、吸着、およびプロトプラスト融合が含まれる。組換え細胞は、単細胞のままであることができ、または成長して組織、臓器、もしくは多細胞生物になることができる。形質転換された本発明のポリヌクレオチド分子は、染色体外に残ることができ、または一体化して、発現されるその能力が保持される様式で、形質転換された(すなわち、組換え)細胞の染色体内の1つもしくは複数の部位になることができる。
【0124】
形質転換するのに適した宿主細胞には、本発明のポリヌクレオチドで形質転換され得る任意の細胞が含まれる。本発明の宿主細胞は、本明細書に記載されるポリペプチドを内因的に(すなわち、天然に)産生することができ、または本明細書に記載される少なくとも1つのポリヌクレオチド分子で形質転換された後そのようなポリペプチドを産生することができる。本発明の宿主細胞は、本明細書に定義される少なくとも1つのタンパク質を産生することができる任意の細胞とすることができ、細菌、真菌(酵母を含む)、寄生虫、線虫、節足動物、動物、および植物の細胞を含む。宿主細胞の例として、サルモネラ(Salmonella)、エシェリキア(Escherichia)、バシラス、リステリア(Listeria)、サッカロミセス(Saccharomyces)、スポドプテラ(Spodoptera)、マイコバクテリア(Mycobacteria)、トリコプルシア(Trichoplusia)、BHK(仔ハムスター腎臓)細胞、MDCK細胞、CRFK細胞、CV−1細胞、COS(例えば、COS−7)細胞、およびベロ細胞が挙げられる。宿主細胞のさらなる例は、大腸菌K−12誘導体を含めた大腸菌;チフス菌(Salmonella typhi);弱毒菌株を含めたネズミチフス菌(Salmonella typhimurium);スポドプテラ・フルギペルダ(Spodoptera frugiperda);イラクサギンウワバ(Trichoplusia ni);および非腫瘍形成性マウス筋原細胞G8細胞(例えば、ATCC CRL 1246)である。有用な酵母細胞は、ピシア属(Pichia sp.)、アスペルギルス属(Aspergillus sp.)およびサッカロミセス属(Saccharomyces sp.)を含む。特に好適な宿主細胞は、細菌細胞、酵母細胞または植物細胞である。
【0125】
組換えDNA技術は、例えば、宿主細胞内でのポリヌクレオチド分子のコピー数、これらのポリヌクレオチド分子が転写される効率、得られる転写物が翻訳される効率、および翻訳後修飾の効率を操作することによって、形質転換ポリヌクレオチド分子の発現を改善するのに使用することができる。本発明のポリヌクレオチド分子の発現を増大させるのに有用な組換え技法として、それだけに限らないが、高コピー数プラスミドへのポリヌクレオチド分子の作動可能な連結、1つまたは複数の宿主細胞染色体中へのポリヌクレオチド分子の組込み、プラスミドへのベクター安定性配列の付加、転写制御シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または修飾、翻訳制御シグナル(例えば、リボソーム結合部位、シャイン−ダルガノ配列)の置換または修飾、宿主細胞のコドン使用に対応するための本発明のポリヌクレオチド分子の修飾、および転写物を不安定化する配列の削除が挙げられる。
【0126】
トランスジェニック植物
本発明を実行する際に使用するのに企図されている植物には、単子葉植物および双子葉植物の両方が含まれる。標的植物には、それだけに限らないが、以下のものが含まれる:シリアル(例えば、コムギ、オオムギ、ライムギ、カラスムギ、米、トウモロコシ、ソルガム、および関連作物);ビート(サトウダイコンおよび飼料用ビート);ナシ状果、核果および柔らかい果物(リンゴ、セイヨウナシ、セイヨウスモモ、モモ、アーモンド、サクランボ、イチゴ、ラズベリー、およびブラックベリー);マメ科植物(マメ、レンズマメ、エンドウマメ、ダイズ);油脂植物(ピーナッツ、セイヨウアブラナ、カラシナ、ケシ、オリーブ、ヒマワリ、ココナツ、ヒマシ油植物、カカオ豆、アメリカホドイモ);キュウリ植物(マロー、キュウリ、メロン);繊維植物(綿、アマ、麻、ジュート);柑橘果実(オレンジ、レモン、グレープフルーツ、マンダリン);野菜(ホウレンソウ、レタス、アスパラガス、キャベツ、ニンジン、タマネギ、トマト、ジャガイモ、パプリカ);クスノキ科(lauraceae)(アボカド、シナモン、樟脳);またはタバコ、木の実、コーヒー、サトウキビ、茶、つる植物、ホップ、芝生、バナナ、などの植物、ならびに天然のゴム植物、ならびに観賞植物(花、低木、広葉樹、および針葉樹などの常緑樹)。本発明の標的植物である、アスペルギルス菌種の感染による影響を頻繁に受ける作物には、これに限定されないが、穀類(トウモロコシ、ソルガム、トウジンビエ、イネ、コムギ)、油糧種子(ピーナッツ、ダイズ、ヒマワリ、ワタ)、香辛料(トウガラシ、クロコショウ、コリアンダー、ウコン、ショウガ)および木の実(アーモンド、ピスタチオ、クルミ、ココナツ)が挙げられる。好ましい実施形態では、植物は、製糖用植物、ワタ、トウモロコシ、ソルガム、パイナップル、クリスマスツリーなどの針葉樹、ユーカリノキ、コムギ、カラスムギ、オオムギ、イネおよびキャノーラから選択される。
【0127】
用語「植物」は、本明細書で名詞として使用する場合、植物全体、例えば、商業用の小麦生産用の田畑で成長する植物などを指す。「植物部分」は、植物構造(例えば、葉、幹)、根、花の器官/構造、種子(胚、胚乳、および種皮を含む)、植物組織(例えば、脈管、組織、基本組織など)、細胞、およびこれらの子孫を指す。
【0128】
本発明に照らして定義されるトランスジェニック植物には、所望の植物または植物器官において本発明の少なくとも1つのポリペプチドを産生させるように、組換え技法を使用して遺伝的に改変された植物(ならびに前記植物の部分および細胞)およびその子孫が含まれる。トランスジェニック植物は、A.Slaterら、Plant Biotechnology−The Genetic Manipulation of Plants、Oxford University Press(2003);ならびにP.ChristouおよびH.Klee、Handbook of Plant Biotechnology、John Wiley and Sons(2004)において一般に記載されているものなどの、当技術分野で既知の技法を使用して作製することができる。
【0129】
「トランスジェニック植物」は、同じ種、種類、または栽培品種の野生型植物において見出されない遺伝子コンストラクト(「導入遺伝子」)を含有する。本明細書で呼ばれる「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって生成または変更され、植物細胞内に導入された遺伝子配列を含む。導入遺伝子は、植物細胞に由来する遺伝子配列を含むことができる。一般に、導入遺伝子は、人間の操作、例えば、形質転換などによって植物中に導入されているが、当業者が認識する任意の方法を使用することができる。
【0130】
好ましい実施形態では、トランスジェニック植物は、その子孫が、所望の表現型を分離しないように導入された各遺伝子およびあらゆる遺伝子(導入遺伝子)についてホモ接合である。トランスジェニック植物は、例えば、ハイブリッド種子から成長したF1子孫内などに導入された導入遺伝子(複数可)についてヘテロ接合とすることもできる。そのような植物は、当技術分野で周知であるハイブリッドの活力(hybrid vigour)などの利点をもたらすことができる。
【0131】
本発明のポリヌクレオチドは、発生のすべての段階の間に、トランスジェニック植物内で構成的に発現され得る。植物または植物器官の使用に応じて、ポリペプチドは、段階特異的な様式で発現され得る。さらに、ポリヌクレオチドは、組織特異的に発現され得る。
【0132】
植物内で対象とするポリペプチドをコードする遺伝子の発現を引き起こすことが知られているか、または見出されている制御配列を、本発明において使用することができる。使用される制御配列の選択は、対象とする標的植物および/または標的器官に依存する。そのような制御配列は、植物または植物ウイルスから得ることができ、または化学的に合成することができる。そのような制御配列は、当業者に周知である。
【0133】
植物細胞の安定なトランスフェクション、またはトランスジェニック植物の確立に適したいくつかのベクターは、例えば、Pouwelsら、Cloning Vectors:A Laboratory Manual(1985、補遺1987);WeissbachおよびWeissbach、Methods for Plant Molecular Biology、Academic Press(1989);ならびにGelvinら、Plant Molecular Biology Manual、Kluwer Academic Publishers(1990)において記載されている。一般に、植物発現ベクターは、5’および3’制御配列の転写制御下の1つまたは複数のクローン化植物遺伝子、および優性選択マーカーを含む。そのような植物発現ベクターは、プロモーター調節領域(例えば、誘導性もしくは構成的な、環境的もしくは発生的に制御された、または細胞もしくは組織特異的発現を制御する調節領域)、転写開始部位、リボソーム結合部位、RNA処理シグナル、転写終止部位、および/またはポリアデニル化シグナルも含有することができる。
【0134】
植物細胞内で活性であるいくつかの構成的プロモーターが記載されている。植物内の構成的発現に適したプロモーターとして、それだけに限らないが、カリフラワーモザイクウイルス(CaMV)35Sプロモーター、ゴマノハグサモザイクウイルス(FMV)35S、サトウキビ桿菌状ウイルスプロモーター、ツユクサ(commelina)黄斑ウイルスプロモーター、リブロース−1,5−ビス−リン酸カルボキシラーゼの小サブユニットに由来する光誘導プロモーター、イネサイトゾルトリオースリン酸イソメラーゼプロモーター、シロイヌナズナ(Arabidopsis)のアデニンホスホリボシルトランスフェラーゼプロモーター、イネアクチン1遺伝子プロモーター、マンノピン合成酵素およびオクトピン合成酵素プロモーター、Adhプロモーター、スクロース合成酵素プロモーター、R遺伝子複合体プロモーター、およびクロロフィルα/β結合タンパク質遺伝子プロモーターが挙げられる。これらのプロモーターは、植物内で発現されたDNAベクターを作り出すのに使用されている;例えば、WO84/02913を参照。これらのプロモーターのすべては、様々な型の植物で発現可能な組換えDNAベクターを作り出すのに使用されている。
【0135】
植物の起源組織、例えば、葉、種子、根、または茎などにおける発現の目的のために、本発明において利用されるプロモーターは、これらの特定の組織において相対的に高い発現を有することが好適である。この目的のために、組織もしくは細胞特異的または組織もしくは細胞で増強される発現を伴う遺伝子についてのいくつかのプロモーターから1つを選択することができる。文献で報告されたそのようなプロモーターの例として、エンドウマメに由来する葉緑体グルタミン合成酵素GS2プロモーター、コムギに由来する葉緑体フルクトース−1,6−ビホスファターゼ(biphosphatase)プロモーター、ジャガイモに由来する核光合成ST−LS1プロモーター、シロイヌナズナ(Arabidopsis thaliana)に由来するセリン/スレオニンキナーゼプロモーター、およびグルコアミラーゼ(CHS)プロモーターが挙げられる。イースタンラーチ(アメリカカラマツ(Larix laricina))に由来するリブロース−1,5−二リン酸カルボキシラーゼプロモーター、マツに由来するCab遺伝子、Cab6についてのプロモーター、コムギに由来するCab−1遺伝子についてのプロモーター、ホウレンソウに由来するCab−1遺伝子についてのプロモーター、イネに由来するCab1R遺伝子についてのプロモーター、トウモロコシ(Zea mays)に由来するピルビン酸オルトリン酸ジキナーゼ(PPDK)プロモーター、タバコLhcb1*2遺伝子についてのプロモーター、シロイヌナズナSuc2スクロース−H30共輸送体プロモーター、ホウレンソウに由来するチラコイド膜タンパク質についてのプロモーター(PsaD、PsaF、PsaE、PC、FNR、AtpC、AtpD、Cab、RbcS)も光合成的に活性な組織において活性であると報告されている。
【0136】
クロロフィルα/β結合タンパク質についての他のプロモーター、例えば、白ガラシ(シナピス・アルバ(Sinapis alba))に由来するLhcB遺伝子およびPsbP遺伝子についてのプロモーターなども本発明において利用することができる。(1)熱、(2)光(例えば、エンドウマメRbcS−3Aプロモーター、トウモロコシRbcSプロモーター);(3)アブシシン酸などのホルモン(4)創傷(例えば、WunI);もしくは(5)化学物質、例えば、メチルジャスミネート(jasminate)、サリチル酸、ステロイドホルモン、アルコール、セーフナー(WO97/06269)などによって調節されるプロモーターを含めた、環境、ホルモン、化学および/または発生シグナルに対する応答において調節される様々な植物遺伝子プロモーターも植物細胞内のRNA結合タンパク質遺伝子の発現のために使用することができ、または(6)器官特異的なプロモーターを使用することも有利となり得る。
【0137】
植物のシンク組織、例えば、ジャガイモ植物の塊茎、トマトの果実、またはダイズ、アブラナ、綿、トウモロコシ、コムギ、イネ、およびオオムギの種子などにおける発現の目的のために、本発明において利用されるプロモーターは、これらの特定の組織において相対的に高い発現を有することが好適である。クラスIパタチンプロモーター、大小サブユニットの両方のジャガイモ塊茎ADPGPP遺伝子についてのプロモーター、スクロース合成酵素プロモーター、22kDのタンパク質複合体およびプロテイナーゼ阻害剤を含めた主要な塊茎タンパク質についてのプロモーター、顆粒結合型デンプン合成酵素遺伝子(GBSS)についてのプロモーター、ならびに他のクラスIおよびIIパタチンプロモーターを含めて、塊茎特異的または塊茎で増強される発現を伴う遺伝子についてのいくつかのプロモーターが知られている。他のプロモーターも、種子または果実などの特定の組織内でタンパク質を発現させるのに使用することができる。β−コングリシニンについてのプロモーター、または他の種子特異的なプロモーター、例えば、ナピンおよびファセオリンプロモーターなどを使用することができる。トウモロコシ胚乳発現について特に好適なプロモーターは、イネに由来するグルテリン遺伝子についてのプロモーター、より具体的には、Osgt−1プロモーターである。コムギにおける発現に適したプロモーターの例には、ADPグルコースピロシンターゼ(pyrosynthase)(ADPGPP)サブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素および脱分枝酵素、胚形成主要タンパク質、グリアジン、ならびにグルテニンについてのプロモーターが含まれる。イネにおけるかかるプロモーターの例には、ADPGPPサブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素、脱分岐酵素、スクロース合成酵素ならびにグルテリンについてのプロモーターが挙げられる。特に好ましいプロモーターは、イネグルテリンについてのプロモーター、すなわちOsgt−1遺伝子である。オオムギについてのかかるプロモーターの例には、ADPGPPサブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素、脱分岐酵素、スクロース合成酵素、ホルデイン、胚グロブリンならびにアリューロン特異的なタンパク質についてのプロモーターが挙げられる。
【0138】
根に特異的なプロモーターも使用することができる。そのようなプロモーターの例は、酸性キチナーゼ遺伝子についてのプロモーターである。根組織における発現は、同定されたCaMV35Sプロモーターの根に特異的なサブドメインを利用することによっても実現することができる。
【0139】
5’非翻訳リーダー配列は、本発明のポリヌクレオチドの異種遺伝子配列を発現するように選択されたプロモーターから得ることができ、必要に応じてmRNAの翻訳を増大するように特異的に改変することができる。導入遺伝子の発現を最適化することの概説については、Kozielら(1996)を参照。5’非翻訳領域は、適当な真核生物遺伝子、植物遺伝子(コムギおよびトウモロコシクロロフィルa/b結合タンパク質遺伝子リーダー)に由来する植物ウイルスRNA(とりわけ、タバコモザイクウイルス、タバコエッチウイルス、トウモロコシ萎縮モザイクウイルス、アルファルファモザイクウイルス)、または合成遺伝子配列から得ることもできる。本発明は、非翻訳領域が、プロモーター配列を伴う5’非翻訳配列に由来するコンストラクトに限定されない。リーダー配列は、無関係のプロモーターまたはコード配列に由来する場合もある。本発明の脈絡において有用なリーダー配列は、トウモロコシHsp70リーダー(US5,362,865およびUS5,859,347)、およびTMVΩ要素を含む。
【0140】
転写の終止は、対象とするポリヌクレオチドにキメラベクター中で作動可能に連結された3’非翻訳DNA配列によって実現される。組換えDNA分子の3’非翻訳領域は、RNAの3’末端にアデニル酸ヌクレオチドの付加を引き起こすように植物内で機能するポリアデニル化シグナルを含有する。3’非翻訳領域は、植物細胞内で発現される様々な遺伝子から得ることができる。ノパリン合成酵素3’非翻訳領域、エンドウマメ小サブユニットRubisco遺伝子に由来する3’非翻訳領域、ダイズ7S種子貯蔵タンパク質遺伝子に由来する3’非翻訳領域は一般に、この能力において使用される。アグロバクテリウム(Agrobacterium)腫瘍誘導(Ti)プラスミド遺伝子のポリアデニル酸シグナルを含有する3’転写非翻訳領域も適している。
【0141】
細胞内に遺伝子を直接送達するための4つの一般的な方法、すなわち、(1)化学的方法(Grahamら、1973);(2)微量注入などの物理的方法(Capecchi、1980);電気穿孔(例えば、WO87/06614、US5,472,869、5,384,253、WO92/09696、およびWO93/21335を参照);および遺伝子銃(例えば、US4,945,050およびUS5,141,131を参照);(3)ウイルスベクター(Clapp、1993;Luら、1993;Eglitisら、1988);ならびに(4)受容体媒介機構(Curielら、1992;Wagnerら、1992)が記載されている。
【0142】
使用することができる加速方法には、例えば、微粒子銃などが含まれる。植物細胞に形質転換核酸分子を送達するための方法の一例は、微粒子銃である。この方法は、Yangら、Particle Bombardment Technology for Gene Transfer、Oxford Press、Oxford、England(1994)によって概説されている。非生物学的粒子(マイクロプロジェクタイル)は、核酸をコーティングし、推進力によって細胞内に送達する。例示的な粒子には、タングステン、金、白金などからなるものが含まれる。微粒子銃が単子葉植物を再現性よく形質転換する有効な手段であることに加えて、微粒子銃の特定の利点は、プロトプラストの分離もアグロバクテリウム感染の感受性も必要とされないことである。加速によってトウモロコシ細胞内にDNAを送達するための方法の例示的な実施形態は、微粒子銃α粒子送達システムであり、これは、DNAでコーティングされた粒子を、ステンレス鋼またはNytexスクリーンなどのスクリーンを通して、懸濁液中で培養されたトウモロコシ細胞で覆われたフィルター表面上に推進するのに使用することができる。本発明とともに使用するのに適した粒子送達システムは、ヘリウム加速PDS−1000/He銃であり、Bio−Rad Laboratoriesから入手可能である。
【0143】
照射のために、懸濁液中の細胞をフィルター上で濃縮することができる。照射される細胞を含有するフィルターは、マイクロプロジェクタイル停止プレートの下に適切な距離をおいて配置される。必要に応じて、1つまたは複数のスクリーンも、銃と照射される細胞との間に配置される。
【0144】
あるいは、未成熟胚または他の標的細胞を、固体培地上に配置することができる。照射される細胞は、マイクロプロジェクタイル停止プレートの下に適切な距離をおいて配置される。必要に応じて、1つまたは複数のスクリーンも、加速デバイスと照射される細胞との間に配置される。本明細書に示した方法を使用することによって、1000またはそれ以上の焦点の、マーカー遺伝子を一過性に発現する細胞を得ることができる。照射して48時間後に外来遺伝子産物を発現する焦点内の細胞数は、1〜10個、平均で1〜3個の範囲であることが多い。
【0145】
照射形質転換において、照射前の培養条件および照射パラメータを最適化することによって、最大数の安定な形質転換体を得ることができる。照射についての物理的および生物学的パラメータの両方が、この技術において重要である。物理的要因は、DNA/マイクロプロジェクタイル沈殿物の操作を伴うもの、またはマクロもしくはマイクロプロジェクタイルの飛行および速度に影響するものである。生物学的要因には、照射前および照射直後の細胞の操作に関与するすべてのステップ、照射に伴う外傷の軽減に役立てるための標的細胞の浸透圧調整、ならびに直線化DNAまたは無傷のスーパーコイルプラスミドなどの形質転換DNAの性質が含まれる。照射前の操作は、未成熟胚の形質転換の成功に特に重要であると考えられている。
【0146】
別の代替の実施形態では、プラスチドを安定に形質転換することができる。高等植物におけるプラスチド形質転換について開示された方法には、選択マーカーを含有するDNAの粒子銃送達、および相同的組換えを通じたプラスチドゲノムへのDNAの標的化が含まれる(US5,451,513、US5,545,818、US5,877,402、US5,932479、およびWO99/05265)。
【0147】
したがって、条件を完全に最適化するための小規模研究において、照射パラメータの様々な態様を調整することが望まれる場合があることが企図されている。物理的パラメータ、例えば、ギャップ距離、飛行距離、組織距離、およびヘリウム圧力などを調整することが特に望まれる場合がある。レシピエント細胞の生理的状態に影響し、したがって形質転換および組込み効率に影響し得る条件を修正することによって外傷悪化要因を最小限にすることもできる。例えば、レシピエント細胞の浸透圧状態、組織水和、および継代培養段階または細胞周期を、最適な形質転換のために調整することができる。他の通常の調整を実施することも、本開示を踏まえると当業者に分かるであろう。
【0148】
アグロバクテリウム媒介トランスファーは、植物細胞内に遺伝子を導入するために広く適用可能なシステムであり、その理由は、DNAを全植物組織内に導入し、それによってプロトプラストから無傷の植物を再生する必要を回避することができるためである。植物細胞内にDNAを導入するために、ベクターを組み込んでいるアグロバクテリウム媒介植物を使用することは、当技術分野で周知である(例えば、US5,177,010、US5,104,310、US5,004,863、US5,159,135を参照)。さらに、T−DNAの組込みは、比較的正確なプロセスであり、ほとんど再配列をもたらさない。トランスファーされるDNAの領域は、ボーダー配列によって画定され、介在DNAは、植物ゲノム内に通常挿入される。
【0149】
最新のアグロバクテリウム形質転換ベクターは、(Kleeら、Plant DNA Infectious Agents、HohnおよびSchell編、Springer−Verlag、New York、179〜203頁(1985))に記載されているように、大腸菌ならびにアグロバクテリウム内で複製することができ、好都合な操作を可能にしている。さらに、アグロバクテリウム媒介遺伝子トランスファーのためのベクターにおける技術的進歩は、ベクター中の遺伝子および制御部位の配置を改善することによって、様々なポリペプチドをコードする遺伝子を発現することができるベクターの構築を促進している。記載されたベクターは、挿入されたポリペプチドコード遺伝子を直接発現させるために、プロモーターおよびポリアデニル化部位に隣接して、好都合なマルチリンカー領域を有し、本目的に適している。さらに、アームドおよびディスアームドTi遺伝子(armed and disarmed Ti gene)を含有するアグロバクテリウムを、形質転換のために使用することができる。アグロバクテリウム媒介形質転換が効率的である植物の種類において、遺伝子トランスファーの容易で明確な性質のために、これが最適の方法である。
【0150】
アグロバクテリウム形質転換法を使用して形成されたトランスジェニック植物は一般に、1つの染色体上に1つの遺伝子座を含有する。そのようなトランスジェニック植物は、付加された遺伝子についてヘミ接合であると呼ぶことができる。より好適なのは、付加された構造遺伝子についてホモ接合であるトランスジェニック植物、すなわち、染色体対の各染色体上の同じ座位に1つの遺伝子という、2つの付加された遺伝子を含有するトランスジェニック植物である。ホモ接合トランスジェニック植物は、1つの付加された遺伝子を含有する独立した分離個体のトランスジェニック植物を有性交配(自家受粉)し、産生された種子のいくつかを発芽させ、対象とする遺伝子について得られた植物を分析することによって得ることができる。
【0151】
2つの異なるトランスジェニック植物を交配することによって、2つの独立して分離している外来遺伝子を含有する子孫を産生させることもできることも理解されるべきである。適切な子孫を自家受粉することにより、両方の外来遺伝子についてホモ接合である植物を産生させることができる。親植物への戻し交配、および非トランスジェニック植物との外交配も、栄養繁殖のように企図されている。様々な形質および作物に一般に使用される他の育種法の記述は、Fehr、Breeding Methods for Cultivar Development、Wilcox J.編、American Society of Agronomy、Madison Wis(1987)に見出すことができる。
【0152】
植物プロトプラストの形質転換は、リン酸カルシウム沈殿、ポリエチレングリコール処理、電気穿孔、およびこれらの処理の組合せに基づく方法を使用して実現することができる。様々な植物の種類にこれらのシステムを適用することは、プロトプラストからその特定の植物系統を再生する能力に依存する。プロトプラストから穀類を再生するための例示的方法が記載されている(Fujimuraら、1985;Toriyamaら、1986;Abdullahら、1986)。
【0153】
細胞形質転換の他の方法も使用することができ、これには、それだけに限らないが、花粉中にDNAを直接トランスファーすること、植物の生殖器中にDNAを直接注射すること、または未成熟胚の細胞内にDNAを直接注射し、その後乾燥された胚を再水和することによる植物中へのDNAの導入が含まれる。
【0154】
1つの植物プロトプラスト形質転換体、または様々な形質転換された外植片から植物を再生し、発生させ、培養することは、当技術分野で周知である(Weissbachら、Methods for Plant Molecular Biology、Academic Press、San Diego、Calif.(1988))。この再生および成長プロセスは一般に、形質転換細胞を選択するステップ、胚発生の通常の段階を介し、根付いた小植物段階を介して個別化した細胞を培養するステップを含む。トランスジェニック胚および種子も同様に再生される。得られたトランスジェニックの根付いた苗条は、土壌などの適切な植物成長培地にその後植えられる。
【0155】
異種の、外来遺伝子を含有する植物の発生または再生は、当技術分野で周知である。再生された植物は、自家受粉することによって、ホモ接合トランスジェニック植物をもたらすことが好ましい。さもなければ、再生された植物から得られる花粉は、農学的に重要な系統の種子から成長した植物と交雑される。反対に、これらの重要な系統の植物からの花粉は、再生された植物に授粉するのに使用される。所望の外因性核酸を含有する本発明のトランスジェニック植物は、当業者に周知の方法を使用して栽培される。
【0156】
主にアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を使用することによって双子葉植物を形質転換し、トランスジェニック植物を得るための方法は、綿(US5,004,863、US5,159,135、US5,518,908);ダイズ(US5,569,834、US5,416,011);アブラナ属(Brassica)(US5,463,174);ピーナッツ(Chengら、1996);およびエンドウマメ(Grantら、1995)について公開されている。
【0157】
外因性核酸を導入することによって植物中に遺伝的変異を導入するため、およびプロトプラストまたは未成熟植物胚から植物を再生するために、コムギおよびオオムギなどの穀類植物を形質転換するための方法は、当技術分野で周知である。例えば、CA2,092,588、AU61781/94、AU667939、US6,100,447、WO97/048814、US5,589,617、US6,541,257およびWO99/14314を参照。トランスジェニックコムギまたはオオムギ植物は、アグロバクテリウム・ツメファシエンス媒介形質転換手順によって産生されることが好ましい。所望の核酸コンストラクトを担持するベクターは、組織培養された植物もしくは外植片の再生可能なコムギ細胞、またはプロトプラストなどの適当な植物系内に導入することができる。
【0158】
再生可能なコムギ細胞は好ましくは、未成熟胚、成熟胚の胚盤、これらの組織または分裂組織から得られるカルスに由来する。
【0159】
トランスジェニック細胞および植物内に導入遺伝子が存在することを確認するために、ポリメラーゼ連鎖反応(PCR)増幅またはサザンブロット分析を、当業者に既知の方法を使用して実施することができる。導入遺伝子の発現産物は、産物の性質に応じて様々な方法のうちのいずれでも検出することができ、ウエスタンブロットおよび酵素アッセイを含む。タンパク質発現を定量化し、様々な植物組織における複製を検出するための1つの特に有用な方法は、GUSなどのレポーター遺伝子を使用することである。トランスジェニック植物が得られたら、これらを成長させることによって、所望の表現型を有する植物組織または植物部分を生成することができる。植物組織または植物部分を回収することができ、かつ/または種子を収集することができる。種子は、所望の特徴を有する組織または部分を伴った追加の植物を成長させるための供給源として機能を果たすことができる。
【0160】
ある実施形態では、本発明のトランスジェニック植物は、US6,369,299に一般的に記載された方法を使用して作製される。
【0161】
トランスジェニック非ヒト動物
「トランスジェニック非ヒト動物」は、同じ種または品種の野生型動物において見出されない遺伝子コンストラクト(「導入遺伝子」)を含有する、ヒト以外の動物を指す。本明細書で呼ばれる「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって生成または変更され、動物細胞内に導入された遺伝子配列を含む。導入遺伝子は、動物細胞に由来する遺伝子配列を含むことができる。一般に、導入遺伝子は、人間の操作、例えば、形質転換などによって動物中に導入されているが、当業者が認識する任意の方法を使用することができる。
【0162】
トランスジェニック動物を作製するための技法は、当技術分野で周知である。この課題に対する有用な一般的な教科書は、Houdebine、Transgenic animals−Generation and Use(Harwood Academic、1997)である。
【0163】
異種のDNAを、例えば、哺乳動物の受精卵子中に導入することができる。例えば、全能性または多能性幹細胞を、微量注入、リン酸カルシウム媒介沈殿、リポソーム融合、レトロウイルス感染または他の手段によって形質転換することができる。次いで形質転換細胞は、胚内に導入され、次いで胚は、発達してトランスジェニック動物になる。非常に好適な方法では、発達中の胚に、所望のDNAを含有するレトロウイルスを感染させ、感染した胚からトランスジェニック動物が生じる。しかし、もっとも好適な方法では、適切なDNAが、好ましくは単細胞段階で胚の前核または細胞質中に同時注入され、胚を発達させて成熟したトランスジェニック動物にする。
【0164】
トランスジェニック動物を作製するのに使用される別の方法は、標準的な方法によって前核段階の卵子中に核酸を微量注入することを伴う。次いで注入された卵子は、培養された後、偽妊娠のレシピエントの輸卵管に移される。
【0165】
トランスジェニック動物は、核トランスファー技術によっても作製することができる。この方法を使用して、ドナー動物に由来する線維芽細胞は、制御配列の制御下で、対象とする結合ドメインまたは結合パートナーについてのコード配列を組み込んでいるプラスミドで安定にトランスフェクトされる。次いで安定なトランスフェクタントは、除核された卵母細胞に融合され、培養され、メスのレシピエントに移される。
【0166】
組成物
本発明の組成物は、「許容可能な担体」とも本明細書で呼ばれる、賦形剤を含む。賦形剤は、処置される動物、植物、植物もしくは動物物質、または環境(土壌および水試料を含む)が耐容性を示すことができる任意の物質とすることができる。そのような賦形剤の例として、水、食塩水、リンガー液、デキストロース溶液、ハンクス液、および他の水性生理学的平衡塩溶液が挙げられる。非水ビヒクル、例えば、固定油、ゴマ油、オレイン酸エチル、または中性脂肪なども使用することができる。他の有用な製剤は、粘度増強剤、例えば、ナトリウムカルボキシメチルセルロース、ソルビトール、またはデキストランなどを含有する懸濁液を含む。賦形剤は、等張性および化学的安定性を増強する物質などの軽微な量の添加剤も含有することができる。緩衝液の例には、リン酸緩衝液、炭酸水素塩緩衝液、およびTris緩衝液が含まれ、一方、保存剤の例には、チメロサール、またはo−クレゾール、ホルマリン、およびベンジルアルコールが含まれる。賦形剤は、組成物、例えば、それだけに限らないが、ポリマー制御放出ビヒクル、生分解性インプラント、リポソーム、細菌、ウイルス、他の細胞、油、エステル、およびグリコールの半減期を増大させるのにも使用することができる。
【0167】
ある実施形態では、本発明のポリペプチドは、固体支持体上に固定される。これにより、s−トリアジンまたはジアジンの加水分解速度および/または程度を増強でき、および/またはポリペプチドの安定性を高めることができる。例えば、ポリペプチドは、ポリウレタン基質上に固定化し(Gordonら、1999)、または適切なリポソーム中にカプセル化することができる(Petrikovicsら、2000aおよびb)。ポリペプチドは、消火において日常的に使用されるものなどのフォームを含む組成物中に組み込むこともできる(LeJeuneら、1998)。当業者に理解されるように、本発明のポリペプチドは、WO00/64539に開示のようにスポンジまたはフォームですぐに使用することができる。本発明に有用な他の固体支持体には、アクリル型構造を有する樹脂、セパビーズEC−EP(Resindion srl−−三菱化学株式会社)およびEupergit C(Rohm−Degussa)などのエポキシ官能基を有する樹脂、またはセパビーズEC−hasおよびEC−EA(Resindion srl−−三菱化学株式会社)などの第1級アミノ基を有する樹脂が挙げられる。いずれの場合も、ポリペプチドは、樹脂と接触させられ、酵素がマトリックスに結合するように、官能基(エポキシド)の高い反応性、またはグルタルアルデヒドなどの二官能性作用剤による樹脂の活性化により固定される。本発明に適した他の樹脂は、ポリスチレン樹脂、巨大網状樹脂およびセパビーズEC−Q1Aなどの塩基性官能基を有する樹脂であり、ポリペプチドは樹脂上に吸収され、次いで二官能性作用剤(グルタルアルデヒド)との架橋結合によって安定化する。
【0168】
ある実施形態では、組成物は、Zn2+および/またはCo2+を含む。別の実施形態では、s−トリアジンまたはジアジンを加水分解するための本発明の方法は、本発明のポリペプチドについての補助因子としてZn2+および/またはCo2+を提供するステップを含む。
【0169】
本発明の一実施形態は、本発明の組成物を、動物、植物、動物もしくは植物物質、または環境(土壌および水試料を含む)中に徐々に放出することができる制御放出製剤である。本明細書で使用する場合、制御放出製剤は、制御放出ビヒクル中に本発明の組成物を含む。適当な制御放出ビヒクルとして、それだけに限らないが、生体適合性ポリマー、他のポリマー基質、カプセル、マイクロカプセル、微粒子、大量瞬時投与製剤、浸透圧ポンプ、拡散デバイス、リポソーム、リポスフェア、および経皮送達システムが挙げられる。好適な制御放出製剤は、生分解性(すなわち、生体内分解性)である。
【0170】
本発明の好適な制御放出製剤は、S−トリアジンまたはジアジンを含む範囲内にある土壌または水中に本発明の組成物を放出することができる。この製剤は好ましくは、約1〜約12カ月の範囲の時間にわたって放出される。本発明の好適な制御放出製剤は、好ましくは少なくとも約1カ月間、より好ましくは少なくとも約3カ月間、さらにより好ましくは少なくとも約6カ月間、さらにより好ましくは少なくとも約9カ月間、さらにより好ましくは少なくとも約12カ月間処理を行うことができる。
【0171】
S−トリアジンまたはジアジンを加水分解するための有効な組成物を生成するのに必要とされる本発明のポリペプチド、ベクター、または宿主細胞などの濃度は、除染される試料の性質、試料中のS−トリアジンまたはジアジンの濃度、ならびに組成物の製剤に依存する。組成物内のポリペプチド、ベクター、または宿主細胞などの有効濃度は、本発明の方法を用いて実験的に容易に求めることができる。
【0172】
本発明の酵素および/またはそれをコードする宿主細胞は、WO2004/112482およびWO2005/26269に一般に記載されているように、コーティング組成物で使用することができる。
【実施例1】
【0173】
増強した活性を有するTrzN突然変異体
方法および材料
トランケートしたpET14bプラスミド(pETcc2)を、TrzNおよびその変異体の発現のために使用した。ユニークなNdeIおよびBamHI部位を用いて、全trzN遺伝子をpETcc2中にクローニングし、大腸菌株BL21λDE3(Novagen)中で発現させた。NdeI/BamHI消化型pETcc2をpETcc2::egfp(図2)から調製し、図2ではライブラリーにおける再連結したベクター(すなわち、青色光下で強く蛍光を発する連結したベクター)の比率の単純なビジュアル表示を提供した。
【0174】
コドンを最適化したTrzN(配列番号2−TrzNco)を、GENEART AG(BioPark、Josef−Engert−Str.11、D−93053 レーゲンブルグ、ドイツ)によって産生した。
【0175】
ランダムな変異誘発を、製品説明書に従い、GeneMorphII(Stratagen)を用いて行った。オリゴヌクレオチドプライマー1および2を、遺伝子の増幅に使用した(表2)。
【0176】
表2に詳述のように、部位飽和変異誘発(Site-saturation mutagenesis)を、プライマー3〜24を用いて、PCR媒介型部位特異的変異誘発によって行った。NNS縮重(Georgescuら、Directed Evolution Library Creation、編:F.H.Arnold、G.Georgiou、Humana Press、Totowa、NJ、2003、pp.75〜89)を、trzN遺伝子の67番目、91番目、131番目、159番目、161番目、243番目、246番目、294番目、335番目および350番目コドンで多様性を生じさせるために使用した(ここでは、Nは任意のヌクレオチドであり、SはGまたはCのいずれかである)。
【0177】
ランダムな部位飽和したライブラリーを、アンピシリン200μg.mL−1、IPTG1μMを補充したLB寒天プレートを用いて選別し、アトラジン1mg.mL−1(90%アトラジンw/w;Gesaprim 900 WG、Syngenta)を浸透させた。TrzN活性を、アトラジン脱塩素化によって評価し、結果として活性TrzNを発現するコロニー付近の培地の清澄化が生じた。活性レベルを、清澄化が起きる速度によって決定した。
【0178】
全配列決定を、Micromon(モナッシュ大学、メルボルン、ビクトリア)によって行った。
【0179】
以前に報告されたように(Shapirら、2005b)、264nMでの吸光度の減少により測定するとき、His6TrzNcc3.2を精製することにより、基質の加水分解速度を測定することによって、基質の範囲を試験した。アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンおよびシアノジン(>99%純度Pestinal標準;Sigma)を、この方法を用いて試験した。His6TrzNcc3.2をアフィニティークロマトグラフィー(HisTrap;GE HealthCare)、続いてサイズ排除クロマトグラフィー(Superdex200;GE HealthCare)を用いて精製した。
【0180】
【表2】
【0181】
結果
反復性ランダム変異誘発
アトラジンのプレート清澄化アッセイを、アトラジンを加水分解的に脱塩素化するBL21λDE3pETcc2::trzNcoの能力を評価するために使用した。30日後に、細菌増殖付近における固体培地の清澄化は観察されなかったため、BL21λDE3pETcc2::trzNcoは、アトラジンのクロロヒドロラーゼ活性レベルは、無かったか検出できなかったと考えられた。trzNcoのランダムな突然変異体を、ローフィデリティのDNAポリメラーゼMutazyme II(GeneMorph II)を用いて作製し、アトラジンに対する加水分解活性について、寒天プレートを含むアトラジンにおいて選別した。trzNcoにおける単一の同義突然変異(T468C)により、156番目のコドンをコードするフェニルアラニンがTTTからTTCに変わった(配列番号3)。突然変異体(TrzN L1、表3)は、37℃で8日後にアトラジンを清澄化する能力がもたらされた。
【0182】
ランダムな変異誘発の次のラウンドについて、TrzN L1を鋳型として使用した(反復1)。27個の突然変異体において、親trzNcoより素早く清澄ゾーンを形成する能力がBL21にもたらされることを発見した(TrzN L1、表3)。清澄ゾーンは、反復1の突然変異体について、3から6日の間で出現した。清澄ゾーンをもっとも素早く形成する15個(TrzN cc1.1、TrzN cc1.3、TrzN cc1.4、TrzN cc1.6、TrzN cc1.8、TrzN cc1.9、TrzN cc1.10、TrzN cc1.11、TrzN cc1.12、TrzN cc1.13、TrzN cc1.14、TrzN cc1.15、TrzN cc1.25、TrzN cc1.26およびTrzN cc1.27)の突然変異体を、ランダム変異誘発の別のラウンドのための鋳型として使用した(反復2)。
【0183】
全体で最良に実行できる突然変異体を選択できるように、反復2の変異誘発PCRからの産物を一緒に選別した。45〜96時間で清澄ゾーンを形成する36個のコロニーを選択した(表3)。反復2の最良の13個の突然変異体(45〜48時間の間で清澄化する;TrzN cc2.1、TrzN cc2.2、TrzN cc2.3、TrzN cc2.4、TrzN cc2.5、TrzN cc2.6、TrzN cc2.7、TrzN cc2.8、TrzN cc2.9、TrzN cc2.10、TrzN cc2.11、TrzN cc2.12およびTrzN cc2.13)を、ランダム変異誘発の3回目の反復のための鋳型として使用した。
【0184】
【表3−1】
【表3−2】
【0185】
再び、全体で最良に実行できる突然変異体を選択できるように、反復2の変異誘発PCRからの産物を一緒に選別した。3回目の反復の13個の突然変異体を選択し、24〜31時間の清澄化時間をもたらすものを選択した。突然変異体におけるヌクレオチドおよびアミノ酸の置換は、表4および5にそれぞれまとめる。単一遺伝子内でもっとも多いヌクレオチド突然変異は、14個であり、任意の変異体酵素において最大7つのアミノ酸置換であった。
【0186】
【表4】
【0187】
【表5−1】
【表5−2】
【0188】
興味深いことに、両方の突然変異が、アミノ酸置換をもたらし、清澄化速度に影響を与えないようであったが、これは、タンパク質のフォールディングおよび安定性に影響する因子に加えて、全体のTrzN活性の転写または翻訳の決定因子が存在することを示唆する。このことは、タンパク質配列は変化させなかったが、むしろF156をコードするのに使用したフェニルアラニンのコドンを(TTTからTTCに)変えた、trzN L1突然変異(T468C)へのtrzNcoの作用によって、明らかに実際に示される。それらが同一産物であるにも関わらず、trzN L1は、明らかにその親遺伝子より優れていた(表3)。
【0189】
部位飽和変異誘発
ランダム変異誘発プログラムの1回目、2回目もしくは3回目の反復の間で、1回より多く変化した、またはランダム変異誘発の2回目および3回目の反復に強く選択された、またはランダム変異誘発の1つのラウンドに存在し、独立した2回目の部位突然変異として、ランダム変異誘発の2回目および3回目の反復の親鋳型において他の突然変異と組み合わせたアミノ酸は、trzNco突然変異体の改良したアトラジンデクロリナーゼ活性の重要な決定因子であると考えられた。ランダム変異誘発によって導入されそうにないアミノ酸との置換を含む、これらの位置の配列スペースをより完全に探索するために、部位飽和ライブラリーをこれらの各部位(コドン67、91、131、159、161、210、243、246、294、335および350;表6)について作成した。
【0190】
【表6】
【0191】
いくつかの部位での別のアミノ酸の広いスペクトルによるアミノ酸置換により、親遺伝子と比較して、清澄化速度が上昇する結果となった一方、他のものは、1つの別のアミノ酸のみへのかなり保存的な変化を許すのみであった。12個(P、N、T、D、V、G、C、S、Q、H、YまたはI)、6個(D、S、E、K、VまたはA)および5個(Y、N、F、KまたはH)の別のアミノ酸に対するL131、D350およびG246のそれぞれの置換により、野生型酵素と比較して増加したデクロリナーゼ活性がもたらされた。A294は、3つの別のアミノ酸(T、SまたはL)によって置換し、より高いアトラジンデクロリナーゼ活性を産生することができた一方、A159およびL243は、2つの別の各アミノ酸(それぞれVまたはT、およびPまたはG)のみによってうまく置換することができた。5つの位置で、1つの可能性のある置換のみが、改良された活性を産生した(Y67F、T91S、S161G、P210AおよびL335M)(表6)。我々はまた、L131P A159VバックグラウンドにおけるD38X部位飽和ライブラリーから100個のコロニーを選別した。我々は、明らかに改良された清澄化の表現型を有する5個のコロニーを得た。かかる表現型をもたらす置換は、D38Nのみであった。
【0192】
変異体Trz Ncc3.2における突然変異の特性評価
SDS−PAGEおよび動力学的解析により、かなり大部分の活性の改良が、増加した発現の結果であることが実際に示された。実際に、発現は、変異誘発の各ラウンドで徐々に増加し、3回目のラウンドで得たもっとも活性の高い突然変異体は、3つのアミノ酸置換(D38N、 L131PおよびA159V)を含んだ(図3)。trzNにおいて多くのサイレント突然変異があったが、これらは、tRNAの利用、mRNAの安定性または翻訳効率への二次構造の影響の変化により、発現の増加に寄与したようである。各アミノ酸変化の溶解性の増加への寄与は、これらをTrzN野生型バックグラウンドに個々に導入することにより、および逆方向突然変異を突然変異体に個々に導入することにより評価した。A159V置換の存在、すなわち、変異誘発の間に導入された第1突然変異により、最終変異体の発現の12.5%(8.4mg/L)に可溶性発現が増強された。A159Vバックグラウンドへの第2置換(L131P)の導入により、最終変異体の発現の42%(28mg/L)に可溶性発現が増強され、L131P、A159Vバックグラウンドへの第3置換(D38N)の導入により、67.2mg/Lに可溶性発現が増加した(図3)。野生型TrzNバックグラウンドにおいて、L131P突然変異は、溶解性においてA159V置換と類似の効果を有した(6.6mg/L収率)。したがって、突然変異を組み合わせることの効果は、相加的というよりむしろ相乗的である。実際に、D38N、L131変異体は、L131P変異体よりかなり低い可溶性であったが、D38N変異体の収率は、野生型TrzNについて以前に報告された収率と類似しており、このことは、A159V置換が存在する場合を除き、D38N置換が、酵素収率にネガティブに影響することを示唆する。
【0193】
基質の範囲
サイズ排除クロマトグラフィーによるTrzN cc3.2の精製により、これは75から150kDaの間の天然の分子量を有することが明らかになったが、このことは、それがホモ二量体またはホモ三量体であることを示唆する。これは、ホモ六量体であるAtzAと対照的であり、TrzNは単量体であるという以前の報告と対照的である(Shapirら、2006)。
【0194】
精製した酵素を、反復3のTrzN変異体の基質範囲を決定するために使用した。TrzN cc3.2は、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンおよびシアノジンを加水分解できたが(表7)、これらは、ハロゲンおよびメチルチオール脱離基、N−エチル、N−イソルプロピル、N−ジエチルならびにN−シアノジメチルメチルアルキル側鎖を有するs−トリアジンを代表する。メチルチオールおよびメトキシ脱離基の化学的性質が高度に類似するため、TrzNcc3.2は、以前に報告された、メトキシ−s−トリアジン(例えば、アトラトン)に対する活性を保持することが期待される(Shapirら、2005b)
【0195】
【表7】
【0196】
宿主およびベクターの範囲
TrzNcc3.2および野生型TrzNの両方についての全体のコード領域を、誘導性の高レベルな発現ベクターpETcc2から、低レベルの恒常的な発現ベクター中に移動した(pCS150、Scottら、2009)。結果のベクターを、大腸菌JM109、DH10βおよびBL21λDE3細胞の形質転換に使用した。結果の6つの菌株を、アトラジン清澄化プレートアッセイを用いて、それらの清澄化速度について試験した。各場合において、TrzNcc3.2発現菌株は、37℃で2日以内に清澄化したが、野生型TrzNを発現する菌株は、37℃で12日後も清澄化しなかった。これは、TrzN発現の改良が、プラスミドまたは菌株のどちらの依存性でもなかったことを実際に示す。
【実施例2】
【0197】
増強した活性を有するTrzN突然変異体の構造
方法および材料
構造解析
データ収集の統計は、他で報告されている(Jacksonら、2006)。単一波長異常回折(SAD)(Dauterら、2002)を用いる金属酵素の活性部位の金属からの相決定は、以前に報告されている(Liuら、2005)。以前の仕事により、非対称性ユニットは、TrzN二量体(Jacksonら、2006)を含むことが示唆されたため、CCP4一式のプログラム(Collaborative−Computational−Research−Project−4、1994)において実行されるように、SHELXD(Schneiderら、2002)を、1.28269Aで収集されたSDAデータを使用する異常差異パターソン合成(anomalous difference Patterson synthesis)において、4つのZn2+イオン部位の位置を決めるのに使用した。MLPHARE(Otwinowski、1991)を、続いて、4つの部位の占有率を精密にし、2.43の相力を用いて初期相を得るために使用した。SHELXEは、密度修正および相の改良のために使用した(Sheldrickら、2002);高い溶媒含量(79%)は、疑いの余地なく相の質に寄与した(Terwilliger、2001)。
【0198】
モデル構築および精密化
初期相を、ARP−wARP(Perrakisら、2001)を用いる自動モデル構築を行うために使用した。これは、38.0%のRfreeを有する初期モデルを作製した。次いで相互作用的なモデル構築のいくつかのラウンドを、COOT(EmsleyおよびCowtan、2004)を用いて実行し、続いてREFMACv5.0(Murshudovら、1997)において実行するように構造の理想化をした後、Rfreeが28.7%になった。抑制した精密化および水分子の追加により、Rfreeは23.5%に減少した。次いでBファクターを、各鎖の残基1〜195、196〜255および256〜271を含む3つの剛体を用いて、TLS精密化(Winnら、2001)の20および10ラウンドにセットし、続いて最大尤度の精密化の3ラウンドにより、Rfreeは20.0%次いで19.6%までさらに低下した。TLSANL(Howlinら、1993)およびANISOANL(Winn、2001)を、TLS精密化によって生成されたデータを解析するために使用した。TLSANLによって生成されたリブレーションテンソル(libration tensor)を、1.5のスケール係数を用いて、RIBBONS(Carson、1991)を用いて可視化した。構造の幾何学的検証を、RAMPAGE(Lovellら、2003)を用いて行い、これによって全残基が、好都合なまたは許容される領域にあったことが示された;PROCHECK(Laskowskiら、1993)では、全立体化学パラメータが、より良いか正常の限界内であることを示し、SFCHECK(Vaguineら、1999)では、0.45A2のLuzattiプロットにより座標において全体のエラーを与えた。
【0199】
結果
TrzNcc3.2の構造を、サルガッソー海の環境試料から、27%同一の構造2PAJを用いて、分子の置換によって解析した(Argawal,R.ら、出版されていない)。正確な解析は、探索モデルの広範な「枝刈り」の後にのみ見出され、さらに次いで二量体の1つの分子のみを見出すことができ、これはプログラムPHASER(McCoyら、2007)を用いる2番目に良いヒットであった。二量体における2つ目の分子を、続いてプログラムMOLREP(VaginおよびTeplyakov、2000)を用いて見出した。53.4%のモデルに関する初期Rfreeは、広範なモデル構築および精密化の後、21.1%に減少した。
【0200】
TrzNの効果的に同一な49.6kDaの2つのサブユニット(図4)を含み、極角において、(θ、φ)=(93、90°)で非結晶性の2回回転軸2倍軸と関連する、結晶性非対称ユニットにおける99.8kDaの1つのホモ二量体が存在する。既知の構造との低い配列類似性にもかかわらず、TrzNcc3.2は、(β/α)8バレルの構造のフォールドを採用し、機能的多様性をもつ大きな金属依存性アミドヒドロラーゼスーパーファミリーに属する。2PAJなどの、タンパク質データバンク(PDB)におけるもっとも類似した構造は、ほとんどが未知の機能である。しかし、機能的注釈がされているもののうち、もっとも近い相対物は、21%配列同一性を有し、2.2Aの主鎖r.m.s.dである、サーモトガ・マリティマ由来のTm0938、5−メチルチオアデノシン/S−アデノシルホモシステイン/アデノシンデアミナーゼ(Hermannら、2007)、19%配列同一性を有し、2.5Aの主鎖r.m.s.dであるヒトグアニンデアミナーゼ(2uz9;Moche,Mら、出版されていない)、および19%配列同位性を有し、2.9Aの主鎖r.m.s.dである、枯草菌由来のイミダゾロンプロピオナーゼである(Yuら、2006)。
【0201】
TrzNの活性部位空洞(図5)は、各サブユニットの触媒ドメインの中央に位置する。基質のアトラジンがドッキングした活性部位アーキテクチャーの図は、図4に示される。ストランド2からのグルタミン残基に加えて、ストランド1(H63、H65)および5(H238)上に位置する、アミドヒドロラーゼモチーフの3つの保存されたヒスチジン、および推定上の水/ヒドロキシド求核試薬は、金属リガンドを構築する。活性部位のZn2+金属イオンは、三方両錐形で配位し、ここでH63、H65およびH238はエクアトリアルなリガンドを含み、Q142および水分子は軸のリガンドを含む。Q129との結合長(3.5A)は、予想したより長いが、この残基の最小の移動によって顕著に短くすることができた。Zn2+金属イオンが、低い占有率(約20%)で結合するときのみ、我々は、完全に占領された活性部位が、より強いQ129−Zn2+相互作用を有し得ることを示唆する。金属の低い占有率が、電子密度に基づくその決定を曖昧とするため、ZnのK−エッジで収集した異常データを、金属酵素において活性部位の金属イオンを同定するのに効果的であることが示されている、Zn2+−TrzNの活性部位のBijvoet差異フーリエマップを計算するために使用した。このマップは、Zn2+が、予想した位置において活性部位で結合することを明白に示した。求核性の水は、アミドヒドロラーゼモチーフの保存された残基であるH274と付加的に水素結合し、ひいてはD300と水素結合して、H−D触媒性二分子を形成することが示される。異なるpH値での動力学的解析により、求核試薬は、溶液中で8.4のpKa値を有する、Zn2+−OH−のさらなる活性化と一致する、およそ8のpKaを有することが示唆される(図9)。
【0202】
一見、金属イオン依存性アミドヒドロラーゼファミリーの他のメンバーの活性部位と類似しているが、1つの大きな違いがある:他の全アミドヒドロラーゼ構造に存在する保存されたアスパラギン酸金属リガンドが、金属イオンリガンドとして働けないスレオニン残基と置き換わっている。アスパラギン酸は、類似しない位置でQ129と明らかに機能的に置き換わっており、これによって金属イオンの三方両錐配位形が回復される。荷電金属イオンリガンドの極性リガンドによる置換は、金属イオン親和性を低下させる可能性がある(図7)。エンテロバクター・アエロゲネス由来の金属ホスホジエステラーゼの活性部位は、この点において情報を与える;活性部位における2つの金属イオンの配位圏は、α部位におけるアスパラギン酸の、β位置におけるアスパラギンによる置換が異なり、これによってβ部位に対する金属イオン親和性の著しい低下が生じることが示された。実際に、TrzNも、金属イオンに対する相対的に弱い親和性を有するようである;過剰なZn2+を増殖培地に添加し、金属キレート剤を、精製または結晶化の間に使用しなかったが、結晶構造は、亜鉛の非常に低い占有率を有する(上記を参照されたい)。
【0203】
Zn2−についての解離定数を、多くのZn2+含有酵素について決定されるものより5から6オーダーの間のマグニチュードで高い、2.6μMであると決定し(図6)、これは、しばしば低いピコモーラー範囲で亜鉛についてKdを有する(炭酸脱水酵素Kd=4pM;グリオキシラーゼI、Kd=27pM;ジペプチジルペプチダーゼIII、Kd=17pM;カルボキシペプチダーゼA、Kd=1.6pMおよびスーパーオキシドジスムターゼ、Kd=10pM。
【0204】
TrzNの基質結合ポケットを特性評価するために、アトラジンを手動で活性部位中にモデリングした(図5および7)。空洞を、4つの区切りに分けることができる;イソプロピルおよびエチル側鎖ポケット、アトラジンの芳香環とπ−πスタッキングにより相互作用する残基、ならびに基質および/または産物と水素結合する残基。金属イオンリガンドH238に加えて、イソプロピル側鎖ポケットは、M82、L86、P131、F132、M163、C198およびY215の側鎖によって形成される。エチル側鎖ポケットは、求核試薬リガンドH274ならびに4つの残基の側鎖P299、D300、M303およびW305によって形成される。ポケットの「ベース」で、W85は、アトラジンの芳香環とπ−πスタッキング相互作用を形成し、これによって結合および遷移状態の間に発生する負電荷の両方を安定化することになる。最後に、E241は、カルボキシル基の酸素原子が、イソプロピル側鎖のNH基から3.3Aであるとき、アトラジンと水素結合を形成するように位置するが、2.9A水素結合によってリンクしたS328−T325二分子は、加水分解によって産生される塩素イオンと相互作用できるようになり、4面体型中間体上に発生する負電荷を安定化する。
【0205】
興味深いことに、ノカルディオイデス菌種株AN3由来の天然TrzN変異体における最近の研究により、アメトリンなどの乏しい脱離基を有する基質に対する触媒活性を消滅させるE241Q突然変異が同定された(Yamazakiら、2008)。本発明者等らは、我々の系におけるこれらの影響を試験し、この突然変異の結果として活性が変化することも発見したが、結合(Km)というよりむしろ主に基質のターンオーバー(kcat)の低下の結果であることを示唆する(表8)。ターンオーバー速度の影響を受けるという観察は、乏しい脱離基を有する基質との低下した活性を有するQ241突然変異体とも一致し、これは、突然変異により、反応への活性化エネルギーを効果的に低下させる酵素の能力が低下されることを示唆する。同じ報告において、位置214および215は、乏しい基質への触媒作用に影響を与えることも確認された。再び我々は、これがkcatの低下によることを示す(表8)。全体的に見て、基質結合性空洞の相補的な性質は、形状および疎水性の両方に関して際立っている。基質/産生成物の水素結合に関わる金属リガンドおよび残基を除き、11/12残基は疎水性であり、同等に疎水的なアトラジン基質の密接でエネルギー的に好都合な結合および脱溶媒和を促進することになるであろう。
【0206】
【表8】
【0207】
活性部位がアトラジンに理想的に適合するようであるが、それはバルク溶媒に完全に閉じ、すなわちアトラジンがこの基質結合ポケットに入ることができる道がないことは興味深い。したがってこれは、立体構造動力学によりが、いくらかの信頼を有もってする基質ターンオーバーに不可欠であることを示唆できる事例である。閉じた活性部位間隙は、図5および7に示し、疎水性残基(L86、M92、L172、Y215、L243、M247、M303およびW305)のネットワークは、活性部位の入口で相互作用し、閉じたそれを効果的に「ジッピング」して閉じる。「ロック」は、ループ3のL172であり、これはネットワークのトップに位置し、定位置に他の残基を効果的に保持することになるであろう。活性部位が開くもっともらしい機構は、特定の可動性ループの尖部に位置するL185に主に関わり、ここで残りのタンパク質に関連する平均的なBファクターが、部分的な占有率/高い可動性と非常に高度に一致する。このループにおける立体構造の変化は、活性部位間隙の2つの側を解開放して、同様の酵素で観察されている「ゆらぎ」の挙動で分離し(Jacksonら、2007)、基質が入り、産生成物が出ていくのを可能にするようである。実際に、このファミリーのメンバーにおける表面ループの立体構造の変動の役割は、最近報告されており、触媒作用(閉じる)および拡散(開く)について最適化した立体構造の基質の間で酵素を切り換えることによって、ターンオーバー速度をモジュールする働きをすることができた。
【0208】
TrzNの構造、基質のドッキングおよび動力学反応速度により、触媒機構を、図8に概説したように提案することができる。反応を4つのステップ(i)基質の結合および求核試薬性の産生、(ii)求核攻撃、(iii)4面体型中間体の分解、(iv)産生成物の放出に分けることができる。触媒作用において重要な役割を果たすようであるTrzNの活性部位に存在する2つの触媒性二分子がある。これらのうちの1つ目、D300−H274二分子は、活性部位の亜鉛金属イオンと協調して、求核性水酸化物の産生に必須のようである。活性部位のZn2+イオンは、結合水のpKaを低下させるルイス酸として働くことになる一方、D300により位置づけられ、安定化したH274は、その脱プロトン化に寄与して求核性水酸化物を形成するためのその脱プロトン化に寄与することになるであろう。基質の結合は、主にW85とのπ−πスタッキング相互作用およびE241との水素結合に関わる。乏しい脱離基に対する活性の損失(Yamazakiら、2008)およびこの残基のQ241への変異誘発におけるkcat数の減少(表8)は、E241との静電気的相互作用が、配向の最適化配向に加えて、基質を活性化する働きができることを示唆する。基質の結合および求核試薬性の産生の後、求核攻撃がC4炭素で起こることになり、非局在性の負電荷が新しいヒドロキシル基、塩素原子、および遷移状態の芳香トリアジン環内にわたって分散した非局在性の負電荷とともに、4面体型中間体の形成を引き起こす(アラインメントは不可能なSN2置換を不可能にする)。この電荷は、ヒドロキシル基でD300−H274二分子、塩素原子でS329−T325二分子、および芳香環でW85とのπ−πスタッキングによって安定化することになるであろう。この中間体の分解により、遊離クロライドに加えて、脱塩素化された産生成物ヒドロキシアトラジンが産生されることになるであろう。産生成物の放出には、次いで酵素における立体構造変化および活性部位間隙の開口が必要になる。
【0209】
結論として、以下の残基:M82、W85、L86、M92、P131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325またはS329を、特異的活性、触媒定数(kcat)、基質特異性(Km)、安定性および/または二次速度定数(kcat/Km)を変えるために変化させることができる。
【実施例3】
【0210】
実地試験
方法および材料
ホモジネートを湯汲むTrzNcc3.2を含むホモジネートの調製
活性TrzNcc3.2を含む清澄化した細菌のホモジネートを、最少培地(10.6g/LのKH2PO4、4g/Lの(NH4)2HPO4、1.7g/Lのクエン酸一水和物、31.3mL/Lのグリセロール)で増殖した、TrzNcc3.2を発現するBL21λDE3の発酵物2リットルから調製した。10mL/LのPTM4塩(0.2g/LのD−ビオチン、2.0g/LのCuSO4.5H2O、0.08g/LのNaI、3.0g/LのMnSO4.H2O、0.2g/LのNa2MoO4、0.02g/Lのホウ酸、0.5g/LのCoCl2.6H2O、7.0g/LのZnC1l2、22.0g/LのFeSO4.7H2O、0.5g/LのCaSO4、1mL/LのH2SO4)をオートクレーブ高圧滅菌した後、0.6g/LのMgSO4を添加した。発酵に、アンピシリン150mg/Lおよびチアミン331mg/Lを補充したグリセロールを与え、IPTG11.9mg/Lで誘導した。発酵により、約240gの湿重量の細胞ペレットが産生された(OD600=122)。細胞をpH6.9のMOPS5.2g/L中で懸濁し、次いでホモジナイザーに3回通し、遠心分離によって清澄化した。清澄化したライセートを、0.22μMのフィルターに通して、インタクトな細胞を除去し、DNaseIをインタクトなDNAの除去に使用した。酵素活性を、Souzaら(1996)により記載されたUV光吸収法およびScottら(2009)に記載された比色法の両方を用いて決定した(アトラジン258±19mg/mgライセート/分)。ホモジネートを−80℃で保存し、必要な時に4℃で解凍した。
【0211】
試験ダムの調製
オーストラリアのクイーンズランドの湿潤/乾燥熱帯のクレア近くのサトウキビ農場(緯度19:48、経度147:14)で、約1.5MLの貯水ダムを、所要量のアトラジン(ヘクタールあたり3.3kg)で前処理した農地の灌漑からの源流で満たした。細菌のホモジネート240gを水20リットル中で懸濁し、貯水ダムの表面にわたって均一に広げることにより散布した。ダムをアトラジン汚染流出水で満たす前、酵素を添加する前、および酵素を添加した後の時間間隔で、ダムをアトラジン含有流出水で満たす前に、1リットルの試料2組を採取した。酵素反応を止めるために試料をすぐに氷上に保存した。氷上に置いて4時間以内に試料を凍結させた。
【0212】
アトラジン濃度の決定
アトラジン濃度を、直接注入投与を用いるように修正した、Lewisら(2009)に記載のLCMSMS法、および以下のLCMS法によるCSIRO Entomologyによって、2つの独立した実験室;Queensland Health Forensic and Scientific Services(QHFSS)で決定した。簡潔には、試料100mLを、HClを用いてpH2.8に酸性化し、次いで試料中のアトラジンを、事前に条件付けしたOasis SPE Max Cartridges(Waters、USA)を用いて、固相抽出によって1000倍濃縮し、(アンモニアを含む)MeOH3mL中に溶出した。試料を続いて乾燥させ、MeOH100μl中で溶解した。試料をHPCLによって分離し、265nmでの吸光度を測定することによってアトラジン濃度についてアッセイし、分析物のピーク領域を、Analystソフトウェアを用いて計算した。増幅させた試料は、10%以内の一致であった。HPLCピークの同一性を、質量スペクトル分析によって確認し、それによってアトラジンイオン216m/zを、Agilent ToF−MSD上に抽出した。
【0213】
結果
オーストラリアのクイーンズランドの湿潤/乾燥熱帯のクレア近くのサトウキビ農場(緯度19:48、経度147:14)で、約1.5MLの貯水ダムを、所要量のアトラジン(ヘクタールあたり3.3kg)で前処理した農地の灌漑からの源流で満たした。TrzNcc3.2を産生する細菌のホモジネート240gを、水20リットル中で懸濁し、貯水ダムの表面にわたって均一に広げることによって手で適用アプライした。ダムをアトラジン汚染流出水で満たす前、酵素を添加する前、および酵素を添加した後の時間間隔で、ダムをアトラジン含有流出水で満たす前に、1リットルの試料2組を採取した。酵素反応を止めるために試料をすぐに氷上に保存した。わずか4時間氷上に置いて4時間以内に後、試料を凍結させた。
【0214】
灌漑の放出された水を採取する前、貯水ダム中の水は、アトラジン8〜12μg/Lを含んだ(データは示さない)。灌漑の放出された水で満たした後、アトラジン濃度は、157〜170μg/Lに上昇した(図10)。酵素の添加後、アトラジンの枯渇速度にずれがあり、これは酵素を貯水ダム中の水と混合した速度にもっとも起因するようであった。このやり方で散布した酵素についての「混合相」の持続時間は、レメディエーションされる水体の、体積や、表面積:体積の割合にほぼ確実に依存し、すなわち、より大きな水体や、小さな表面積:体積の割合を有する水体はより長期の混合相を必要とするであろう。
【0215】
混合相の間のずれにもかかわらず、酵素の添加は、添加後、最初の4時間に、アトラジンの濃度における>90%枯渇を導いた。この結果は、トリアジンについてのTrzNベースのバイオレメディエーションが、技術的に実行可能であることを示す。
【0216】
広く記載した本発明の精神または範囲から逸脱することなく、特定の実施形態において示した本発明に、多数の変形および/または改変を行うことができることが、当業者によって理解されるであろう。したがって、本実施形態は、あらゆる点で例示的であり、限定的ではないと見なされるべきである。
【0217】
本願は、US61/094,044からの優先権を主張し、その内容全体は、参照により本明細書に組み込まれている。
【0218】
本明細書で論じ、かつ/または参照したすべての刊行物は、その全体が本明細書に組み込まれている。
【0219】
本明細書に含めた文献、行為、物質、デバイス、物品などの任意の考察は、単に本発明についての背景を提供する目的のためである。任意またはすべてのこれらの事柄は、従来技術の基礎の一部を形成する、またはこれが本願の各請求項の優先日前に存在した場合、本発明に関連した分野における共通の一般的な知見であったことを認めるものと解釈されるべきでない。
【0220】
(参考文献)
【表9−1】
【表9−2】
【0221】
配列表への鍵配列番号1−野生型TrzNのアミノ酸配列
配列番号2−TrzNをコードする、コドンを最適化したオープンリーディングフレーム(TrzNco)
配列番号3−TTTからTTCへの156番目のコドン変化を有するTrzNをコードする、コドンを最適化したオープンリーディングフレーム(Trz L1)
配列番号4−野生型TrzNをコードするオープンリーディングフレーム
配列番号5から30−オリゴヌクレオチドプライマー
【技術分野】
【0001】
本発明は、アトラジンなどのS−トリアジンおよびジアジンを分解するためのポリペプチドに関する。また、これらのポリペプチドをコードするポリヌクレオチドも提供する。本発明は、S−トリアジンおよびジアジンのバイオレメディエーションにおける、これらのポリヌクレオチドおよびポリペプチドの使用にも関する。
【背景技術】
【0002】
最近の集約農業の実行は、トリアジン除草剤などの効果的な化学的ペストコントロール剤の使用によって促進される。例えば、アトラジン(6−クロロ−N2−エチル−N4−イソプロイル−1,3,5−トリアジン−2,4−ジアミン)は、非常に効果的な発芽前および後のトリアジン除草剤であり、1958年に最初に導入されてから、広葉雑草種のコントロール用に広範に使用されてきた(Tomlin、2006)。
【0003】
環境的に意味のある濃度のアトラジンは、因果関係として脊椎動物種において内分泌機能異常に関連し(例えば、アフリカツメガエルの男性性徴消失(demasculination))(Hayesら、2002、2003および2006)、アトラジンが発癌性である可能性があることが示唆されている(Huff、2002;HuffおよびSass、2007)。さらに、それらの幅広い特異性のため、アトラジンおよび関連のトリアジン除草剤は、非標的の光合成種へのそれらの毒性作用により、環境の損害を引き起こす可能性を有する。
【0004】
アトラジンは、環境中で移動性および持続性の両方である。アトラジンの環境的半減期は、4および57週間の間と見積もられており(Belluckら、1991)、アトラジンは、いくつかの国において、地表水および地下水の両方で検出されている(ThurmanおよびMeyer、1996;van der Meer、2006;Gavrilescu、2005)。
【0005】
いくつかの遺伝子/酵素系が、炭素および窒素源として、トリアジン農薬の異化を可能にする原核生物において進化している。もっとも完全に特性評価されたこれらの経路は、シュードモナス菌種ADPからもともと単離された(Mandelbaumら、1995;de Souzaら、1995)、伝播性pADP1プラスミド(Martinezら、2001)に由来するatzABCDEF遺伝子によりコードされる。アトラジンおよびシマジン(6−クロロ−N2,N4−ジエチル−1,3,5−トリアジン−2,4−ジアミン)(de Souzaら、1996)は、atzA、atzBおよびatzCによりコードされるアミドヒドロラーゼファミリー酵素によって、連続的に脱塩素化および脱アルキル化され(de Souzaら、1996;Boundy−Millsら、1997;Sadowskyら、1998)、シアヌル酸を産生し、これは、次いでatzD、atzEおよびatzFによってコードされる、経路中の残りのヒドロラーゼによって、アンモニアおよび二酸化炭素にミネラル化される(Frucheyら、2003;Chengら、2005;Shapirら、2005a)。
【0006】
AtzABCDEFアトラジン分解異化経路のAtzA酵素は、わずか25.4%同一であるにもかかわらず、TrzNトリアジン分解酵素と頻繁に置き換えられ(Sajjaphanら、2004)、これはAtzAと重複する活性を有する(Shapirら、2005b)。TrzNは、亜鉛依存性アミドヒドロラーゼファミリー酵素であり(Shapirら、2006)、トリアジン化合物から塩化物、フッ化物、S−メチル、S(O)−メチルおよびシアノ基の加水分解の置換にを担う(Shapirら、2005b)。これは、TrzNが、幅広くs−トリアジンを標的にする一方、AtzAは、ハロゲン化したs−トリアジンを解毒することのみに使用できることを意味する。TrzNは、クロロ−s−トリアジン(例えば、アトラジン、プロパジンおよびシマジン)、メチルオキシ−s−トリアジン(例えば、アトラトン、シメトンおよびプロメトン)およびメチルチオ−s−トリアジン(例えば、アメトリン、プロメトリンおよびシメトリン)除草剤を標的にする。
【0007】
TrzNはまた、AtzAの5秒−1と比較して、同程度のAtzA2.1秒−1の触媒定数を有するが、アトラジンについてははるかにより低いKmを有することが報告されている(AtzAについて100μMと比較して20μM)(Shapirら、2006)。したがって、AtzAは、3.3×104のアトラジンについてKcat/Kmを有するが、TrzNは、アトラジンについて1×105のKcat/Kmを有し、TrzNは、AtzAより触媒作用が効率的な酵素であることを実際に示す。
【0008】
しかし、AtzAと違い、TrzNは、大腸菌などの異種宿主において有意量で発現させるのは難しいことがわかっている。TrzNを分子シャペロンGroELとともに共発現させたとき、10mg.mL−1より少ない最大収率を大腸菌から得たが(Shapirら、2006)、シャペロンの非存在ではわずか560μg.mL−1の最大収率であった(Shapirら、2005b)。
【0009】
バイオレメディエーションは、潜在的に有害な残留農薬の環境への影響を改善するための新興のアプローチである(Alcaldeら、2007)。成功したバイオレメディエーション戦略の1つは、酵素的バイオレメディエーションのものであり、ここでは単離または半精製した酵素を、農薬の毒性を大幅に低下させるようなやり方で、毒性農薬を異化または修飾するために使用する(Paralesら、2002;Sutherlandら、2004)。酵素的バイオレメディエーションは、生微生物の使用よりも多くの利点を有する;GM生物またはインタクトなDNAの環境中への放出があり、使用する酵素は一般的に素早く(ほんの数時間の適用時間を必要とする)、適用後の制限された予測可能な持続性を有する(Alcaldeら、2007)。
【0010】
しかし、バイオレメディエーションに採用する酵素の要件は、いくらか厳しく、高い触媒活性、低いKm、拡散性の補助因子への非依存、および環境条件(pH、温度、塩濃度など)の範囲に対して一般的に頑強なタンパク質が必要とされる。酵素はまた、大腸菌などの典型的な発酵生物中で、高い可溶性の活性タンパク質として発現しなければならず、この点においてTrzNは不適当である。
【発明の概要】
【発明が解決しようとする課題】
【0011】
アトラジンの潜在的に大きな環境への足跡が心配されるが、農業においてその継続的な使用が所望される。したがって、環境のダメージに対し、アトラジンおよび他のs−トリアジンならびにジアジンの潜在力を除去するか低下させるためのさらなる方法が必要である。
【課題を解決するための手段】
【0012】
本発明者らは、TrzNをコードするポリヌクレオチド、または強化した特性を有するその変異体を同定した。
【0013】
第1の態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解するポリペプチドをコードする、単離された、かつ/または外因性のポリヌクレオチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリヌクレオチドを提供する。
【0014】
好ましい実施形態では、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが、可溶性の生物学的活性形態で産生される。
【0015】
さらに特に好ましい実施形態では、ポリヌクレオチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードする。別の実施形態では、ポリヌクレオチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含むポリペプチドをさらにコードする。
【0016】
好ましい実施形態では、ポリヌクレオチドは、以下のヌクレオチドの置換の1つもしくは複数、またはそこに対応するヌクレオチド位置での置換を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む;T5C、C39A、C76A、C84A、T87C、C101A、T108A、T108A、G112A、A127G、C135T、A157T、C165T、G168A、C180T、C189T、A200T、C207T、G210A、G225T、A228G、C229T、T240C、A250C、C268A、G270A、A271T、T273A、C279T、A296G、A302G、A303G、A314G、C315A、T317C、T320C、A326C、A333G、T336C、C346T、G357A、A367G、C372T、C375A、C381T、T384C、C391A、C391G、C391T、T392C、T392A、T392G、G393C、T399C、C410T、C411A、C411T、A414G、A418C、T423C、T426A、A432T、C438T、C449G、C454T、T466C、T468C、T471C、C474T、G475A、C476T、A481G、C483T、G489A、G489T、T498C、T531A、A537G、A540G、A545G、T546G、G548A、T555C、T555C、C564T、G567A、G567A、G568A、G569A、G573C、T579C、A584T、G589A、A600G、T618C、T618C、T627C、C628G、G630A、C633T、G637A、T639C、T639C、A654G、G660A、G660T、T663C、C675A、G681T、C686T、C690A、C696T、G705A、A723G、C727G、T728C、T728G、G729C、G736A、G737C、G737A、G737T、T738G、T738C、T738C、G745A、G753C、G768A、T774A、C807A、T840A、A843G、A852T、A855G、C867T、T879C、G880A、G880T、G880C、C881T、G882T、C885T、G897A、T900C、T906A、A928G、A938T、T941C、C957A、T959A、C972T、T978A、C981T、C993T、C999T、C1003A、C1003T、T1011C、G1048A、G1048T、G1048C、A1049T、A1049G、G1053A、A1059G、A1086G、G1094A、T1101C、T1101G、C1128T、A1152G、G1176T、C1186A、C1186T、T1196C、C1203T、G1221A、C1223T、C1236T、G1248T、G1270A、C1278T、T1286A、T1305C、G1309A、C1321T、A1326G、C1329T、C1329T、C1332T、C1344A、C1351AおよびG1353T。
【0017】
ある実施形態では、ポリヌクレオチドは、以下のアミノ酸の置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する、配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;
I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【0018】
さらなる実施形態では、ポリヌクレオチドは、以下のアミノ酸の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する、配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;M82、W85、L86、M92、L131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325およびS329。
【0019】
特に好ましい別の実施形態では、ポリヌクレオチドは、配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含む。
【0020】
さらに好ましい実施形態では、ポリヌクレオチドは、
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置に、アスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置に、スレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含むポリペプチドをコードする。
【0021】
別の実施形態では、ポリヌクレオチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0022】
さらに別の実施形態では、ポリヌクレオチドは、以下のヌクレオチド置換もしくは置換グループの1つ、またはそこに対応するヌクレオチド位置(複数可)での置換(複数可)を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む;
i)T468C、
ii)T468C、A938T、
iii)A200T、G210A、T468C、
iv)T468C、C476T、G753C、
v)T468C、C476T、T728C、
vi)T468C、1048T、
vii)T384C、T468C、G569A、G681T、
viii)T468C、G475A、
ix)C279T、T468C、C1223T、C1329T、
x)C76A、T468C、A481G、
xi)C39A、C101A、T468C、T639C、G737C、G1048T、
xii)C410T、A418C、T468C、A600G、
xiii)T468C、G705A、C1003A、
xiv)T468C、G573C、
xv)T468C、C474T、C628G、C1278T、
xvi)T399C、T468C、G880A、T900C、
xvii)T468C、C1236T、
xviii)T87C、A367G、T468C、
xix)T468C、G1176T、
xx)C135T、T468C、C1344A、
xxi)T468C、A852T、
xxii)T468C、T738C、
xxiii)C454T、T468C、
xxiv)A200T、T468C、G1309A、
xxv)T468C、G489T、G745A、
xxvi)C410T、T468C、
xxvii)T468C、G736A、
xxviii)G225T、C268A、A414G、T468C、T627C、C1321T、
xxix)A432T、T468C、T471C、C476T、T728C、G1053A、C1351A、
xxx)A303G、C449G、T468C、C476T、C686T、C690A、T728C、C1128T、
xxxi)A200T、G210A、C372T、T468C、A654G、C1003A、
xxxii)C180T、A200T、A302G、C375A、T399C、C411T、T468C、A540G、G880A、T900C、
xxxiii)C229T、T468C、G705A、C1003A、
xxxiv)T317C、T468C、A481G、T531A、G753C、T906A、T978A、C1003A、A1326G、
xxxv)A127G、T320C、T468C、C476T、G753C、G1048T、
xxxvi)A157T、C410T、A418C、T468C、A545G、G568A、A600G、C628G、G630A、G1048T、C1332T、
xxxvii)T468C、C476T、A723G、C1003A、G1048T、C1329T、
xxxviii)C84A、T399C、T468C、C483T、G880A、T900C、G1048A、
xxxix)T468C、T498C、C628G、C885T、A1086G、G1270A、
xl)T384C、T468C、C476T,G569A、G753C、A1152G、
xli)T336C、T468C、C476T、A537G、C564T、
xlii)A228G、T240C、T468C、C628G、G880A、G1048T、G1094A、
xliii)T273A、A367G、C381T、T399C、T468C、A481G、G880A、T900C、
xliv)A200T、C207T、G210A、T468C、C476T、G1048T、A1059G、
xlv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1236T、
xlvi)A200T、G210A、C346T、T468C、C476T、T579C、T728C、C1278T、
xlvii)C438T、T468C、C476T、A855G、
xlviii)G168A、T426A、T468C、C476T、C628G、C1203T、T1305C、
xlix)T468C、C476T、T663C、C1236T、
l)T384C、T468C、C476T、T728C、A928G、C1003A、C1186A、
li)T5C、C315A、T468C、C476T、G589A、G681T、T728C、G897A、C1003A、
lii)C279T、T468C、C476T、C1003A、
liii)A250C、A314G、T468C、C476T、
liv)A200T、G210A、T468C、T555C、G880A、T900C、
lv)T468C、C476T、T546G、C696T、C1003A、G1048T、
lvi)A200T、G210A、T468C、C476T、T728C、T1101G、
lvii)G112A、C165T、T468C、C476T、T618C、G753C、C999T、T1101C、
lviii)T423C、T468C、C476T、G489A、A584T、G753C、G1048T、
lix)T466C、T468C、C474T、C628G、G1048T、
lx)T108A、A200T、G210A、G357A、T468C、G1048T、A1059G、C1278T
lxi)T468C、C476T、G567A、G753C、G1048T、
lxii)A200T、G210A、A296G、T468C、C476T、G637A、T728C、C1003A、
lxiii)C189T、A326C、T468C、C476T、T941C、T959A、A1059G、C1186T、T1196C、G1248T、T1286A、
lxiv)T468C、C476T、C1003A、
lxv)A200T、G210A、C279T、T468C、C476T、T879C、C1003A、G1048T、
lxvi)G112A、C165T、T392C、T468C、C476T、T618C、G660A、C675A、G753C、C807A、C993T、C999T、T1101C、T1305C、
lxvii)A271T、T392C、T468C、C476T、G753C、G880A、G1048T、G1094A、C1186A、G1221A、
lxviii)A228G、T240C、T468C、G548A、C628G、G1048T、C1332T、
lxix)T108A、A200T、G210A、T423C、T468C、C476T、G567A、G1048T、
lxx)T384C、T468C、C476T、C628G、C867T、G880A、T900C、C981T、G1048A、A1326G、
lxxi)A200T、C207T、G210A、T468C、C476T、T840A、T900C、G1048A、
lxxii)T468C、C476T、C633T、G660T、A723G、C1003A、G1048T、C1329T、
lxxiii)A228G、T240C、G270A、T468C、C628G、880A、G1048T、
lxxiv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1003T、C1236T、
lxxv)A228G、T240C、T468C、C628G、G880A、C957A、C1003A、または
lxxvi)A200T、G210A、C411A、T468C、T555C、T639C、A654G、G768A、T774A、C972T、C1003A、T1011C、G1353T。
【0023】
ある実施形態では、細菌細胞中で少なくとも2回、より好ましくは少なくとも5回発現させると、ポリペプチドの量は、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも産生される。
【0024】
さらなる実施形態では、細菌細胞中で少なくとも2回、より好ましくは5回発現させると、可溶性の生物学的活性ポリペプチドの量は、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも産生される。
【0025】
代替的な実施形態では、大腸菌株BL21λDE3中で発現させ、アンピシリン200μg.mL−1、IPTG1μMを補充したLB寒天プレート上で培養し、アトラジン1mg.mL−1(90%アトラジンw/w)を浸透させるとき、コロニー付近の培地の清澄化は、培養から約8日以内、より好ましくは約6日以内、さらにより好ましくは約2日以内に検出することができる。かかる条件下のポリヌクレオチドの発現は、実施例の部でさらに詳細に記載する。
【0026】
好ましい実施形態では、ポリペプチドは、配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍、より好ましくは少なくとも5倍、さらにより好ましくは7倍のアトラジン加水分解活性を有する。
【0027】
好ましい実施形態では、ポリペプチドは、配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍、より好ましくは少なくとも5倍のシマジン加水分解活性を有する。
【0028】
本発明のポリヌクレオチドによりコードされるポリペプチドによって加水分解できるs−トリアジンの例には、これに限定されないが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンが挙げられる。
【0029】
細菌細胞は、本発明のポリヌクレオチドによってコードされるポリペプチドを産生できる任意の細胞であってよい。好ましい実施形態では、細菌細胞は、大腸菌である。適した大腸菌株の例には、これに限定されないが、BL21λDE3(ATCC寄託番号PTA−2657)、JM109およびDH10βが挙げられる。
【0030】
さらに好ましい実施形態では、ポリヌクレオチドは、細胞中でポリヌクレオチドの発現を導くことができるプロモーターに作動可能に連結されている。
【0031】
別の実施形態では、ポリヌクレオチドは、少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質をコードする。少なくとも1つの他のポリペプチドは、例えば、本発明のポリペプチドの安定性を強化するポリペプチド、細菌細胞もしくは酵母細胞などの細胞からの融合タンパク質の分泌を促すポリペプチド、または融合タンパク質の精製を補助するポリペプチドであってよい。
【0032】
別の態様では、本発明は、本発明のポリヌクレオチドを含むベクターを提供する。
【0033】
また、本発明のポリヌクレオチド、および/または本発明のベクターを含む宿主細胞も提供する。
【0034】
ある実施形態では、宿主細胞は、シャペロンをコードする外因性ポリヌクレオチドをさらに含む。
【0035】
本発明の宿主細胞の例には、これに限定されないが、細菌細胞、酵母細胞または植物細胞が挙げられる。
【0036】
さらなる態様では、本発明は、本発明の少なくとも1つの細胞を含むトランスジェニック植物を提供する。
【0037】
さらに別の態様では、本発明は、本発明の少なくとも1つの細胞を含む、トランスジェニック非ヒト動物を提供する。
【0038】
さらなる態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解する、実質的に精製された、および/または組換えポリペプチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリペプチドを提供する。
【0039】
好ましい実施形態では、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが、可溶性の生物学的活性形態で産生させる。
【0040】
さらに特に好ましい実施形態では、ポリペプチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含む。別の実施形態では、ポリペプチドはさらに、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含む。
【0041】
本発明のポリペプチドによって加水分解できるs−トリアジンの例には、これに限定されないが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンが挙げられる。
【0042】
ある実施形態では、ポリペプチドは、少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質である。少なくとも1つの他のポリペプチドは、例えば、本発明のポリペプチドの安定性を強化するポリペプチド、細菌細胞もしくは酵母細胞などの細胞からの融合タンパク質の分泌を促すポリペプチド、または融合タンパク質の精製を補助するポリペプチドであってよい。
【0043】
別の実施形態では、ポリペプチドは、固体支持体に固定化されている。
【0044】
別の態様では、本発明は、本発明の宿主細胞の抽出物、本発明の植物および/または本発明の動物を提供し、抽出物は、本発明のポリペプチドを含む。
【0045】
さらなる態様では、本発明は、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、および/または本発明の抽出物、および1つもしくは複数の許容される担体を含む組成物を提供する。
【0046】
さらなる態様では、本発明は、s−トリアジンまたはジアジンを加水分解するための方法であって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物と、s−トリアジンまたはジアジンを接触させるステップを含む方法を提供する。
【0047】
ある実施形態では、土壌、水、生体物質またはそれらの組合せからなる群から選択される試料。
【0048】
別の態様では、本発明は、対象においてs−トリアジンまたはジアジンより引き起こされた毒性を処理する方法であって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物を対象に投与するステップを含む方法を提供する。
【0049】
別の態様では、本発明は、対象においてs−トリアジンまたはジアジンにより引き起こされた毒性の処理用の薬剤を製造するための、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明のポリペプチド、本発明の抽出物、および/または本発明の組成物の使用を提供する。
【0050】
別の態様では、本発明は、s−トリアジンおよび/またはジアジンを加水分解できるポリペプチドを産生する方法であって、ポリペプチドをコードするポリヌクレオチドの発現を可能にする条件下で、前記ポリペプチドをコードする本発明の宿主細胞、または前記ポリペプチドをコードする本発明のベクターを培養するステップと、発現したポリペプチドを回収するステップとを含む方法を提供する。
【0051】
なおさらなる態様では、本発明は、宿主細胞を検出するための方法であって、
i)細胞または細胞集団を、細胞(複数可)によるポリヌクレオチドの取込みを可能にする条件下で、本発明のポリヌクレオチドと接触させるステップと、
ii)ステップi)からの細胞またはその子孫細胞を、s−トリアジンまたはジアジンにさらすことにより宿主細胞を選択するステップと
を含む方法を提供する。
【0052】
ある実施形態では、ポリヌクレオチドは、本発明のポリペプチドをコードする。
【0053】
ある実施形態では、ポリヌクレオチドは、本発明のポリヌクレオチドを含む第1オープンリーディングフレーム、および本発明のポリヌクレオチドを含まない第2オープンリーディングフレームを含む。
【0054】
一実施形態では、第2オープンリーディングフレームは、ポリペプチドをコードする。別の実施形態では、第2オープンリーディングフレームは、翻訳されていないポリヌクレオチドをコードする。両場合において、第2オープンリーディングフレームは、適したプロモーターに作動可能に連結されていることが好ましい。
【0055】
好ましくは、翻訳されていないポリヌクレオチドは、触媒性核酸、dsRNA分子またはアンチセンス分子をコードする。
【0056】
好ましい実施形態では、細胞は植物細胞である。
【0057】
さらなる態様では、本発明は、s−トリアジンまたはジアジンを加水分解するためのキットであって、本発明のポリヌクレオチド、本発明のベクター、本発明の宿主細胞、本発明によるポリペプチド、本発明の抽出物、および/または本発明の組成物を含むキットを提供する。
【0058】
さらなる態様では、本発明は、本発明のポリペプチドの結晶を提供する。
【0059】
別の態様では、本発明は、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを設計する方法であって、本発明の結晶の原子座標を使用して、s−トリアジンまたはジアジンが、候補ポリペプチドと結合する能力をコンピュータによって評価するステップと、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを選択するステップとを含む方法を提供する。
【0060】
明らかになるように、本発明の一態様の好ましい特質および特徴は、本発明の他の多くの態様に適用可能である。
【0061】
本明細書を通して、用語「含む(comprise)」、または「含む(comprises)」または「含んでいる(comprising)」などの変形は、述べられた要素、整数もしくはステップ、または要素、整数もしくはステップのグループを含有することを意味するが、他の任意の要素、整数もしくはステップ、または要素、整数もしくはステップのグループを排除しないことを理解されよう。
【0062】
本発明を、以下の非限定的実施例によって、かつ添付の図面を参照して以下に説明する。
【図面の簡単な説明】
【0063】
【図1】本発明の酵素によって分解される様々なs−トリアジンの構造を示す図である。
【図2】pETcc2::egfpの概略図である。
【図3】野生型TrzN、TrzN cc3.2および中間突然変異型の部分的な精製を示す図である。野生型TrzNに関する値は、5リットルから2.8mgを産生するShapirら(2005)から取る。分子量マーカーを含んだが(レーンM)、それらの分子量(kDa)を示す。TrzNおよびその変異体の位置は、矢印で示す。TrzNおよびその変異体の同一の精製から得た同等量(5μl)を、各レーンに添加した。
【図4】TrzN構造の概略図および絵である。TrzNは、99.6kDaのホモ二量体を形成する。各単量体は、2つのドメイン:βサンドイッチドメインおよびβ/α8バレルドメインに分割される。β/α8バレルは、上側の面上にいくつかのループの挿入を有し、これは、活性部位の入口を修飾するのに役立ち、またはさらに二量体の界面を形成する。
【図5】TrzNcc3.2の基質結合ポケットを構成する残基を示す概略図である。アトラジン、Zn2+中心およびヒドロキシルイオン(OH−)を中央に示す。基質結合ポケットを含むアミノ酸の同一性および配列位置を示す。
【図6】Zn2+を有するApo−TrzNcc3.2の飽和を示すグラフである。最大半量の活性は、ZnCl22.6μMで回復した。
【図7】活性部位のアミノ酸残基およびアミノ酸残基に配位する金属の概略図である。アトラジン、Zn2+およびヒドロキシルイオン(OH−)を中央に示し、関連残基の同一性および配列位置を示す。
【図8】提案されたTrzNの反応機構を説明する概略図である。
【図9】TrzNとアメトリンの速度定数(kcat)、および反応緩衝液のpHの間の関係を示すグラフである。
【図10】TrzNcc3.2による実地試験の結果を示すグラフである。TrzNcc3.2を添加して10.5時間後の1.5MLのホールディングダム(holding dam)中のアトラジンの枯渇。QHFSS(四角)およびCSIRO Entomology(丸)によって解析した試料。
【発明を実施するための形態】
【0064】
一般的な技法および定義
他に特に定義されない限り、本明細書で使用されるすべての技術的および科学的用語は、当業者(例えば、細胞培養、バイオレメディエーション、分子遺伝学、免疫学、免疫組織化学、タンパク質化学、および生化学における)によって一般に理解されるのと同じ意味を有すると解釈されるものとする。
【0065】
他に指定されない限り、本発明において利用される組換えタンパク質、細胞培養、および免疫学的技法は、当業者に周知の標準的な手順である。そのような技法は、出典における文献、例えば、J.Perbal、A Practical Guide to Molecular Cloning、John Wiley and Sons(1984);J.Sambrookら、Molecular Cloning:A Laboratory Manual、Cold Spring Harbour Laboratory Press(1989);T.A.Brown(編)、Essential Molecular Biology:A Practical Approach、1巻および2巻、IRL Press(1991);D.M.GloverおよびB.D.Hames(編)、DNA Cloning:A Practical Approach、1〜4巻、IRL Press(1995および1996);およびF.M.Ausubelら(編)、Current Protocols in Molecular Biology、Greene Pub.Associates and Wiley−Interscience(1988、現在までのすべての最新版を含む);Ed HarlowおよびDavid Lane(編)Antibodies:A Laboratory Manual、Cold Spring Harbour Laboratory、(1988);ならびにJ.E.Coliganら(編)、Current Protocols in Immunology、John Wiley&Sons(現在までのすべての最新版を含む)などにわたって記載および説明されている。
【0066】
本明細書で使用するとき、用語「加水分解する」、「加水分解している」、「加水分解活性」およびその変形は、化学結合の加水分解を触媒する本発明のポリペプチドの能力をいう。好ましい実施形態では、本発明の酵素は、以下のものの1つまたは複数;デハロゲナーゼ(例えば、クロロヒドロラーゼ、フルロヒドロラーゼ、ハロヒドロラーゼ、加水分解性デコルリナーゼ(decholrinase)、加水分解性デフルロリナーゼおよび/または加水分解性デハロゲナーゼ)、メトキシヒドロラーゼおよびメチルチオヒドロラーゼである。好ましい実施形態では、酵素活性の産物が、s−トリアジンまたはジアジン基質と比べて、例えば、哺乳動物および/または魚類への毒性が弱くなり、かつ/または安定性が低くなるように、ポリペプチドは、s−トリアジンまたはジアジンを「分解する」。
【0067】
本明細書で使用するとき、用語「より高いs−トリアジンおよび/またはジアジン加水分解活性」とは、s−トリアジンまたはジアジンに関して、より高い特異的活性、触媒定数(kcat)、基質特異性(Km)および/または二次速度定数(kcat/Km)、または配列番号1として提供されるアミノ酸配列を含むポリペプチドよりも高い安定性を有する本発明のポリペプチドをいう。特異的活性は、実施例の概説のように決定することができる。
【0068】
本明細書で使用するとき、表現「そこに対応するアミノ酸位置に」および「アミノ酸番号に対応する位置に」とは、周囲のアミノ酸と比較して、相対的なアミノ酸位置をいう。例えば、いくつかの実施形態では、本発明のポリペプチドは、例えば、配列番号1に対して整列させたとき、アミノ酸の相対的な位置付けを変える、欠失または置換の突然変異を有し得る。ある実施形態では、ポリペプチドは、指定された残基番号に規定されたアミノ酸を含む。
【0069】
同様に、表現「そこに対応するヌクレオチド位置に」および「ヌクレオチド番号に対応する位置に」とは、周囲のヌクレオチドと比較して、相対的なヌクレオチドの位置をいう。例えば、いくつかの実施形態では、本発明のポリヌクレオチドは、例えば、配列番号2または配列番号4に対して整列させたとき、ヌクレオチドの相対的な位置付けを変える、欠失または置換の突然変異を有し得る。ある実施形態では、ポリヌクレオチドは、指定されたヌクレオチド番号に規定されたヌクレオチドを含む。
【0070】
本明細書で使用するとき、用語「処理している」、「処理する」または「処理」は、s−トリアジンまたはジアジンによって引き起こされる毒性の少なくとも1つの症状を和らげるまたは排除するのに十分な、処理効果のある量の本発明のポリペプチド、またはそれをコードするポリヌクレオチドを投与することを含む。
【0071】
用語「生体物質」は、本明細書において、生物起源の任意の産物を含むもっとも広範な意味で使用される。かかる産物には、これに限定されないが、ヒトのための食料産物および動物の餌が挙げられる。この産物には、水およびミルクなどの液体食材、ならびにヨーグルトなどの半流動性の食材を含む液体媒体が挙げられる。本発明はまた、固体食材、特に動物の餌にまで及ぶ。ある実施形態では、生体物質は、これに限定されないが、サトウキビ、キャノーラ種子、コムギ種子、オオムギ種子、ソルガム種子、イネ、トウモロコシ、パイナップルまたはワタ種子などの植物物質であることが好ましい。
【0072】
本明細書で使用するとき、用語「抽出物」とは、本発明の宿主細胞、植物またはトランスジェニック非ヒト動物の任意の部分をいう。この部分は、植物種子などの実体全部であってよく、または少なくとも部分的な均質化および/または精製によって得ることができる。この用語は、宿主細胞から分泌される部分を含み、したがって培養上澄み液を包含する。
【0073】
本明細書で使用するとき、用語「シャペロン」とは、細胞からのタンパク質の排出を変えるために、他のタンパク質が適切なフォールディングまたはアンフォールディングを成し遂げるのを補助する機能を有するタンパク質をいう。シャペロンは当技術分野でよく知られており、これに限定されないが、トリガー因子(TF)などのリボソーム結合性タンパク質;Hsp70、DnaK、Hsp40、DnaJ、GrpEなどのシャペロンのHsp7Oファミリー、およびGroEL、GroES、Hsp60、Hsp10などのシャペロンのシャペロニンファミリーが挙げられる。非限定的な例として、大腸菌由来のシャペロンGroELは、シャペロンの熱ショックタンパク質60(Hsp60)クラスのメンバーであり、大腸菌のGroEオペロン由来のGroESと一緒に発現する。GroELは、凝集しやすいポリペプチド中間体の濃度、および経路から外れる凝集率を減少させるアンフォールドのタンパク質に結合することによって、タンパク質のフォールディング反応を補助し、これによって未変性の立体構造に分割するのを助ける。コシャペロニンGroEL、ならびにATP、K+およびMg2+などの補助因子は、GroEL媒介型ポリペプチドのフォールディング反応の収率をさらに増加させることが知られている。したがって、特定の実施形態では、当業者は、シャペロン媒介型(または補助型)タンパク質フォールディングを改良または強化するために、当技術分野で既知の成分、補助因子、追加のシャペロンタンパク質を含めよう。
GroELをコードするのに使用できるベクターの例には、これに限定されないが、dnaK−dnaJ−grpE−groES−groELを含むpG−KJE8、groES−groELを含むpGro7、およびgroES−groEL−tigを含むpG−Tf2が挙げられる(Nishiharaら、1998および2000)。
【0074】
s−トリアジンおよびジアジン
本明細書で使用するとき、「s−トリアジン」は、3つの炭素原子および3つの窒素原子からなる6員環複素環式芳香族環を有する化学構造を含む有機化合物である。トリアジン環中の原子は、ベンゼン環における原子と類似している。本発明の酵素によって加水分解される(分解される)s−トリアジンのタイプの例には、これに限定されないが、クロロ−s−トリアジン、フルオロ−s−トリアジン、メチルチオ−s−トリアジンおよびメチルオキシ−s−トリアジンが挙げられる。本発明の酵素によって加水分解される(分解される)いくつかのs−トリアジンの化学構造は、図1に示される。好ましい実施形態では、s−トリアジンは、次の構造を有する;
2−R1−4−R2−6−R3−1,3,5−トリアジン、式中R1が、Cl、Fl、OCH3、SCH3、S(O)CH3またはN3であってよく、R2またはR3が、OCH3、NHCH2CH2OH、NH(CH2)2CH3、NH(CH2)3CH3、NHCH2CH(CH3)2、NHCH(CH3)CH2CH3、NHC(CH3)2CN、NHC(CH3)3、NH2またはOHであってよい。
【0075】
塩素化(クロロ)s−トリアジンは、少なくとも1つの塩化物を含む。塩素化s−トリアジンの例には、これに限定されないが、アトラジン(6−クロロ−N−エチル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)(図1参照)、クロラジン(6−クロロ−N,N,N’,N’−テトラエチル−1,3,5−トリアジン−2,4−ジアミン)、シアナジン(2−[[4−クロロ−6−(エチルアミノ)−1,3,5−トリアジン−2−イル]アミノ]−2−メチルプロパンニトリル)、シプラジン(6−クロロ−N−シクロプロピル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、エグリナジン(N−[4−クロロ−6−(エチルアミノ)−1,3,5−トリアジン−2−イル]グリシン)、イパジン(6−クロロ−N,N−ジエチル−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、メソプラジン(6−クロロ−N−(3−メトキシプロピル)−N’−(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、プロシアジン(2−[[4−クロロ−6−(シクロプロピルアミノ)−1,3,5−トリアジン−2−イル]アミノ]−2−メチルプロパンニトリル)、プログリナジンN−[4−クロロ−6−[(1−メチルエチル)アミノ]−1,3,5−トリアジン−2−イル]グリシン)、プロパジン(6−クロロ−N,N’−ビス(1−メチルエチル)−1,3,5−トリアジン−2,4−ジアミン)、セブチルアジン(6−クロロ−N−エチル−N’−(1−メチルプロピル)−1,3,5−トリアジン−2,4−ジアミン、シマジン(6−クロロ−N,N’−ジエチル−1,3,5−トリアジン−2,4−ジアミン)、テルブチルアジン(6−クロロ−N−(1,1−ジメチルエチル)−N’−エチル−1,3,5−トリアジン−2,4−ジアミン)およびトリエタジン(6−クロロ−N,N,N’−トリエチル−1,3,5−トリアジン−2,4−ジアミン)、ならびに(デスエチル(desethyl)アトラジンおよびデスイソプロピル(desisopropyl)アトラジンなどのアトラジン脱アルキル化を含む)クロロ−s−トリアジン脱アルキル化の産物が挙げられる。
【0076】
メチルチオ−s−トリアジンは、少なくとも1つのチオール基を含む。メチルチオ−s−トリアジンの例には、これに限定されないが、アメトリン(N2−エチル−N4−イソプロピル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)、プロメトリン(N2,N4−ジイソプロピル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)およびシメトリン(N2,N4−ジエチル−6−メチルチオ−1,3,5−トリアジン−2,4−ジアミン)が挙げられる。
【0077】
メトキシ−s−トリアジンは、少なくとも1つのメトキシ基を含む。メトキシ−s−トリアジンの例には、これに限定されないが、アトラトン(N2−エチル−N4−イソプロピル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)、シメトン(N2,N4−ジエチル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)およびプロメトン(N2,N4−ジイソプロピル−6−メトキシ−1,3,5−トリアジン−2,4−ジアミン)が挙げられる。
【0078】
フルオロ−s−トリアジンは、少なくとも1つのフッ素を含む。フルオロ−s−トリアジンの例は、フルオルアトラジン(2−フルオロ−4−N−エチルアミノ−6−N−イソプロピルアミノ−1,3,5−トリアジン)である。
【0079】
ある実施形態では、s−トリアジンは、少なくとも1つのN−エチル、N−イソプロピル、N−ジエチルおよび/またはN−シアノジメチルメチルアルキル側鎖を有する。
【0080】
好ましい実施形態では、ジアジンは、次の構造を有する;−2−R1−4−R2−6−R3−1,3−ピリミジンであり、式中R1が、Cl、Fl、OCH3、SCH3、S(O)CH3またはN3であってよく、R2またはR3が、OCH3、NHCH2CH2OH、NH(CH2)2CH3、NH(CH2)3CH3、NHCH2CH(CH3)2、NHCH(CH3)CH2CH3、NHC(CH3)2CN、NHC(CH3)3、NH2またはOHであってよい。ジアジンのタイプの例には、これに限定されないが、クロロ−s−ジアジンおよびフルオロ−s−ジアジンが挙げられる。除草剤であるジアジンの例は、ブロマシル(5−ブロモ−3−sec−ブチル−6−メチルウラシル)である。
【0081】
ポリペプチド
「実質的に精製された」または「精製された」とは、1つまたは複数の脂質、核酸、他のポリペプチド、またはポリペプチドがその天然状態で伴っている他の混入分子から分離されたポリペプチドを意味する。実質的に精製されたポリペプチドは、ポリペプチドが天然に伴っている他の成分を、少なくとも60%、より好ましくは少なくとも75%、より好ましくは少なくとも90%含まないことが好適である。しかし現在、本発明のポリペプチドが自然に存在する証拠はない。
【0082】
ポリペプチドとの関連において用語「組換え体」は、ポリペプチドの天然状態と比較して変化した量、または変化した速度で細胞によって、または無細胞発現系において産生されるときのポリペプチドを指す。一実施形態では、細胞は、ポリペプチドを天然に産生しない細胞である。しかし、細胞は、産生されるポリペプチドの量の変化、好ましくは増加を引き起こす非内因性遺伝子を含む細胞であり得る。本発明の組換えポリペプチドには、これが産生される形質転換(組換え)細胞または無細胞発現系の他の成分から分離されていないポリペプチド、および少なくともいくつかの他の成分から引き続いて精製される、そのような細胞または無細胞系で産生されるポリペプチドが含まれる。
【0083】
用語「ポリペプチド」および「タンパク質」は一般に、互換的に使用され、非アミノ酸基の付加によって修飾されていても、されていなくてもよい1本のポリペプチド鎖を指す。そのようなポリペプチド鎖は、他のポリペプチドもしくはタンパク質、または補助因子などの他の分子と付随することができることが理解される。用語「タンパク質」および「ポリペプチド」は、本明細書で使用する場合、本明細書に記載されるポリペプチドの変異体、突然変異体、生物学的活性断片、修飾体、類似体および/または誘導体も含む。
【0084】
ポリペプチドの%同一性は、ギャップ生成ペナルティー=5、およびギャップ伸長ペナルティー=0.3を用いてGAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)によって決定される。クエリー配列は、長さが少なくとも25アミノ酸であり、ギャップ分析は、少なくとも25アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも50アミノ酸であり、ギャップ分析は、少なくとも50アミノ酸の領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも100アミノ酸であり、ギャップ分析は、少なくとも100アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、クエリー配列は、長さが少なくとも250アミノ酸であり、ギャップ分析は、少なくとも250アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、クエリー配列は、長さが少なくとも400アミノ酸であり、ギャップ分析は、少なくとも400アミノ酸の領域にわたって2つの配列を整列させる。さらにより好ましくは、ギャップ分析は、それらの全長にわたって2つの配列を整列させる。
【0085】
本明細書で使用する場合、「生物学的活性断片」は、全長ポリペプチドの規定された活性を維持する、本明細書に記載されるようなポリペプチドの部分である。生物学的活性断片は、これらが規定された活性を維持する限り任意のサイズとすることができる。好ましくは、生物学的活性断片は、長さが少なくとも100、より好ましくは少なくとも400のアミノ酸である。
【0086】
好ましい実施形態は、「可溶性の生物学的活性形態」で産生されるポリペプチドに関する。当業者が理解するように、これは、細胞中で発現するときに不溶性、したがって、不活性の形態では存在しないポリペプチドをいう。さらに、生物学的活性とは、s−トリアジンおよび/またはジアジンを加水分解する能力を意味する。
【0087】
定義されたポリペプチドに関して、上記に提供したものより高い%同一性の数値は、好適な実施形態を包含することが理解されよう。したがって適用可能な場合、最低%同一性の数値を踏まえると、ポリペプチドは、該当する指定された配列番号と少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%、より好ましくは少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一であるアミノ酸配列を含むことが好適である。
【0088】
本明細書に記載されるポリペプチドのアミノ酸配列突然変異体は、本明細書に定義される核酸中に適当なヌクレオチドの変化を導入することによって、または所望のポリペプチドのインビトロ合成によって調製することができる。そのような突然変異体は、例えば、アミノ酸配列内の残基の欠失、挿入、または置換を含む。最終的なコンストラクトに到達するために、欠失、挿入および置換の組合せを行うことができ、ただし最終的なポリペプチド生成物は、所望の特徴を有するという条件がある。
【0089】
突然変異体(変化した)ポリペプチドは、当技術分野で既知の任意の技法を使用して調製することができる。例えば、本明細書に記載されるポリヌクレオチドは、インビトロ突然変異生成に付すことができる。そのようなインビトロ突然変異生成技法は、適当なベクターにポリヌクレオチドをサブクローニングするステップ、大腸菌(E. coli)XL−1赤色(Stratagene)などの「ミューテーター」菌株にベクターを形質転換するステップ、および適当な数を生成するために形質転換細菌を増殖させるステップを含むことができる。別の例では、本発明のポリヌクレオチドは、Harayama(1998)によって広く記載されたDNAシャッフリング技法に付される。突然変異/変化したDNAに由来する生成物は、これらが、活性の増強および/または基質特異性の変化などの所望の表現型を付与することができるかどうかを判定するために、本明細書に記載される技法を使用して容易にスクリーニングすることができる。
【0090】
アミノ酸配列突然変異体の設計において、突然変異部位の位置および突然変異の性質は、改変される特性(複数可)に依存する。突然変異の部位は、例えば、(1)実現される結果に応じて、最初に保存的なアミノ酸選択物で、次いでより急進的な選択物で置換し、(2)標的残基を欠失させ、または(3)位置づけられた部位に隣接して他の残基を挿入することによって個々に、または連続的に改変することができる。
【0091】
アミノ酸配列の欠失は一般に、約1〜15個の残基、より好ましくは約1〜10個の残基、および典型的には約1〜5個の連続した残基の範囲である。
【0092】
置換突然変異体は、取り出されるポリペプチド分子中の少なくとも1つのアミノ酸残基、およびその場所に挿入される異なる残基を有する。置換突然変異生成にとってもっとも対象となる部位は、機能に対して重要であると同定された部位を含む。対象とする他の部位は、様々な菌株または種から得られる特定の残基が同一であるものである。これらの位置は、生物活性とって重要となり得る。これらの部位、特に、3つの他のまったく同様に保存された部位の配列内に入るものは、相対的に保存された様式で置換されることが好ましい。そのような同類置換を表1に示す。
【0093】
好ましい実施形態では、突然変異体/変異体ポリペプチドは、本明細書で具体的に定義されたポリペプチドと比較したとき、1または2または3または4つの保存アミノ酸変化を有する。保存アミノ酸変化の詳細は、表1に示される。
【0094】
好ましい実施形態では、ポリペプチドは、以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【0095】
【表1】
【0096】
さらなる実施形態では、ポリペプチドは、以下のアミノ酸の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む;M82、W85、L86、M92、L131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325およびS329。これらのアミノ酸の1つまたは複数は、ポリペプチドの特異的活性、触媒定数(kcat)、基質特異性(Km)、安定性および/または二次速度定数(kcat/Km)を変えるように変化させることができる。
【0097】
さらに好ましい実施形態では、ポリペプチドは、
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンまたはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンまたはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンまたはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンまたはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含む。
【0098】
ある実施形態では、ポリペプチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含む;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0099】
より好ましくは、ポリペプチドは、以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換を有する配列番号1に提供されるアミノ酸配列を含む;
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【0100】
特に好ましい実施形態では、ポリペプチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含み、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが、可溶性の生物学的活性形態で産生される。その上、この実施形態では、ポリペプチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンをさらに含むことが好ましい。
【0101】
好ましくは、他に規定されなければ、所与のアミノ酸位置で、ポリペプチドは、配列番号1として提供されるポリペプチドの対応する位置で見られるようなアミノ酸を含む。
【0102】
指定された部位のアミノ酸が、表1に提供されたアミノ酸置換と矛盾するとき、指定されたアミノ酸が好ましい。
【0103】
好ましい実施形態では、ポリペプチドは、本発明の2つの別々のポリペプチド鎖の二量体である。ポリペプチドは、ホモ二量体またはヘテロ二量体であってよい。ヘテロ二量体に関しては、2つのポリペプチド鎖は、少なくとも90%、より好ましくは少なくとも95%、より好ましくは少なくとも97%、より好ましくは少なくとも99%同一であることが好ましい。
【0104】
さらに好ましい実施形態では、ポリペプチドは、Zn2+またはCo2+と結合している。
【0105】
その上、さらに突然変異体を設計するとき、当業者は、TrzNcc3.2の構造に関する実施例2に提供される情報を使用することができる。
【0106】
さらに、必要に応じて、非天然アミノ酸または化学的アミノ酸類似体を、本明細書に記載されるポリペプチド中への置換または付加として導入することができる。そのようなアミノ酸として、一般に、それだけに限らないが、一般的なアミノ酸のD異性体、2,4−ジアミノ酪酸、α−アミノイソ酪酸、4−アミノ酪酸、2−アミノ酪酸、6−アミノヘキサン酸、2−アミノイソ酪酸、3−アミノプロピオン酸、オルニチン、ノルロイシン、ノルバリン、ヒドロキシプロリン、サルコシン、シトルリン、ホモシトルリン、システイン酸、t−ブチルグリシン、t−ブチルアラニン、フェニルグリシン、シクロヘキシルアラニン、β−アラニン、フルオロ−アミノ酸、デザイナーアミノ酸、例えば、β−メチルアミノ酸、Cα−メチルアミノ酸、Nα−メチルアミノ酸など、およびアミノ酸類似体が挙げられる。
【0107】
やはり本発明の範囲内に含まれるのは、例えば、ビオチン化、ベンジル化、グリコシル化、アセチル化、リン酸化、アミド化、既知の保護基/ブロッキング基による誘導体化、タンパク質分解切断、抗体分子または他の細胞リガンドへの連結などによって、合成中、または合成後に差次的に改変される本発明のポリペプチドである。これらの改変は、ポリペプチドの安定性および/または生物活性を増大させるのに機能を果たすことができる。
【0108】
本明細書に記載されるポリペプチドは、天然ポリペプチドの生成および回収、組換えポリペプチドの生成および回収、ならびにポリペプチドの化学合成を含めた様々な方法で生成することができる。一実施形態では、本発明の単離ポリペプチドは、ポリペプチドを生成するのに有効な条件下で、ポリペプチドを発現することができる細胞を培養し、ポリペプチドを回収することによって生成される。培養するのに好適な細胞は、本発明の組換え細胞である。有効な培養条件は、それだけに限らないが、ポリペプチド生成を可能にする有効培地、バイオリアクター、温度、pH、および酸素条件を含む。有効培地は、本発明のポリペプチドを生成するために細胞が培養される任意の培地を指す。そのような培地は一般に、同化できる炭素、窒素、およびリン酸源を有する水媒質、ならびに適切な塩、ミネラル、金属、およびビタミンなどの他の栄養分を含む。本発明の細胞は、従来の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿、およびペトリ皿において培養することができる。培養は、組換え細胞にとって適切な温度、pH、および酸素含量で実施することができる。そのような培養条件は、当業者の専門知識内である。
【0109】
ある実施形態では、本発明のポリペプチドは、細胞からのポリペプチドの分泌を導くことができるシグナル配列を含む。非常に多くのかかるシグナル配列を単離しており、これはNおよびC末端シグナル配列を含む。原核生物および真核生物のN末端シグナル配列は類似しており、真核生物のN末端シグナル配列は、細菌において分泌配列として機能できることが示されている。かかるN末端シグナル配列の例は、よく研究されている配列である、細菌のβ−ラクタマーゼシグナル配列であり、ポリペプチドの外部環境への分泌を促進するために幅広く使用されている。C末端シグナル配列の例は、大腸菌のヘモリジンA(hlyA)シグナル配列である。シグナル配列のさらなる例には、これに限定しないが、アエロリジン、アルカリホスファターゼ遺伝子(phoA)、キチナーゼ、エンドキチナーゼ、α−ヘモリジン、MIpB、プルラナーゼ、YopsおよびTATシグナルペプチドが挙げられる。
【0110】
ポリヌクレオチド
一本鎖または二本鎖の、センスもしくはアンチセンス配向または両方の組合せ、dsRNAまたはその他でのDNA、RNA、またはこれらの組合せを含めた「単離ポリヌクレオチド」とは、これが、その天然状態で付随または連結されているポリヌクレオチド配列から少なくとも部分的に分離されたポリヌクレオチドを意味する。好ましくは、単離ポリヌクレオチドは、これが天然に付随している他の成分を、少なくとも60%、好ましくは少なくとも75%、もっとも好ましくは少なくとも90%含まない。さらに、用語「ポリヌクレオチド」は、用語「核酸」と本明細書で互換的に使用される。
【0111】
ポリヌクレオチドとの関連で、用語「外因性の」は、ポリヌクレオチドの天然状態と比較して変化した量で、細胞内または無細胞発現系内に存在するときのポリヌクレオチドを指す。一実施形態では、細胞は、ポリヌクレオチドを天然に含まない細胞である。しかし、細胞は、コードされるポリペプチドの変化した、好ましくは増加した産生量をもたらしている非内因性ポリヌクレオチドを含む細胞であり得る。本発明の外因性ポリヌクレオチドは、これが存在する形質転換(組換え)細胞または無細胞発現系の他の成分から分離されていないポリヌクレオチド、および少なくともいくつかの他の成分から引き続いて精製される、そのような細胞または無細胞系で産生されるポリヌクレオチドが含まれる。
【0112】
ポリヌクレオチドの%同一性は、ギャップ生成ペナルティー=5、およびギャップ伸長ペナルティー=0.3を用いてGAP(NeedlemanおよびWunsch、1970)分析(GCGプログラム)によって決定される。他に述べられていない限り、クエリー配列は、長さが少なくとも45ヌクレオチドであり、ギャップ分析は、少なくとも45ヌクレオチドの領域にわたって2つの配列を整列させる。好ましくは、クエリー配列は、長さが少なくとも150ヌクレオチドであり、ギャップ分析は、少なくとも150ヌクレオチドの領域にわたって2つの配列を整列させる。より好ましくは、クエリー配列は、長さが少なくとも300ヌクレオチドであり、ギャップ分析は、少なくとも300ヌクレオチドの領域にわたって2つの配列を整列させる。さらにより好ましくは、ギャップ分析は、2つの配列の全長にわたって2つの配列を整列させる。
【0113】
定義されたポリヌクレオチドに関して、上記に提供したものより高い%同一性の数値は、好適な実施形態を包含することが理解されよう。したがって適用可能な場合、最低%同一性の数値を踏まえると、本発明のポリヌクレオチドは、該当する指定された配列番号と少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも80%で、少なくとも85%、より好ましくは少なくとも90%、より好ましくは少なくとも91%、より好ましくは少なくとも92%、より好ましくは少なくとも93%、より好ましくは少なくとも94%、より好ましくは少なくとも95%、より好ましくは少なくとも96%、より好ましくは少なくとも97%、より好ましくは少なくとも98%、より好ましくは少なくとも99%、より好ましくは少なくとも99.1%、より好ましくは少なくとも99.2%、より好ましくは少なくとも99.3%、より好ましくは少なくとも99.4%、より好ましくは少なくとも99.5%、より好ましくは少なくとも99.6%、より好ましくは少なくとも99.7%、より好ましくは少なくとも99.8%、さらにより好ましくは少なくとも99.9%同一である配列を含むことが好適である。
【0114】
本発明はまた、ストリンジェントな条件下で、配列番号2および/または配列番号4をコードするポリヌクレオチドにハイブリダイズするポリヌクレオチドであって、本発明のポリペプチドをコードし、および/または本明細書で規定されるような置換(複数可)を含むポリヌクレオチドにも関する。用語「ストリンジェントなハイブリダイゼーションの条件」または「ストリンジェントな条件」などは、本明細書で使用するとき、ポリヌクレオチドまたはオリゴヌクレオチドの長さとともに様々なハイブリダイゼーションの温度を含む、当技術分野で精通しているパラメータをいう。核酸ハイブリダイゼーションのパラメータは、かかる方法を集めたSambrookら(上記)およびAusubelら(上記)の文献で見つけることができる。例えば、ストリンジェントなハイブリダイゼーションの条件とは、本明細書で使用するとき、ハイブリダイゼーション緩衝液(3.5xSSC、0.02%フィコール、0.02%ポリビニルピロリドン、0.02%ウシ血清アルブミン、2.5mMのNaH2PO4(pH7)、0.5%SDS、2mMのEDTA)中で65℃でのハイブリダイゼーション、および各洗浄ステップは約30分で、65℃で0.2xSSC、0.1%SDS中で2回洗浄することをいう場合がある。
【0115】
特に好ましい実施形態では、ポリヌクレオチドは、配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含み、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが産生される。
【0116】
さらに好ましい実施形態では、ポリヌクレオチドは、配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードし、細菌細胞中で発現するとき、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合より、多くのポリペプチドが、可溶性の生物学的活性形態で産生される。その上、この実施形態では、ポリヌクレオチドは、i)配列番号1のアミノ酸番号38に対応する位置にアスパラギン、およびii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンまたはイソロイシンを含むポリペプチドをさらにコードすることが好ましい。
【0117】
本発明のポリヌクレオチドは、ここで提供される分子と比較したとき、1つまたは複数の突然変異を有することができ、この突然変異は、ヌクレオチド残基の欠失、挿入、または置換である。突然変異体は、天然に存在しても(すなわち、天然源から単離された)、合成であってもよい(例えば、核酸への部位特異的な突然変異生成を実施することによる)。
【0118】
通常、ポリヌクレオチドの単量体は、ホスホジエステル結合またはその類似体によって連結される。ホスホジエステル結合の類似体として、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホラニリデート、およびホスホラミデートが挙げられる。
【0119】
組換えベクター
本発明の一実施形態には、組換えベクターが含まれ、これは、宿主細胞内にポリヌクレオチド分子を送達することができる任意のベクター中に挿入された、少なくとも1つの本発明の単離/外因性ポリヌクレオチドを含む。そのようなベクターは、異種のポリヌクレオチド配列、すなわち、本発明のポリヌクレオチド分子に隣接して天然に見出されず、好ましくはポリヌクレオチド分子(複数可)が由来した種以外の種に由来するポリヌクレオチド配列を含有する。ベクターは、RNAであってもDNAであってもよく、原核生物であっても真核生物であってもよく、一般にトランスポゾン(US5,792,294に記載されているような)、ウイルス、またはプラスミドである。
【0120】
ある型の組換えベクターは、発現ベクターに作動可能に連結したポリヌクレオチド(複数可)を含む。作動可能に連結された、という語句は、分子が、宿主細胞内に形質転換されたとき発現され得るような様式での、発現ベクター内へのポリヌクレオチド分子の挿入を指す。本明細書で使用する場合、発現ベクターは、宿主細胞を形質転換し、指定されたポリヌクレオチド分子の発現を生じさせることができるDNAまたはRNAベクターである。発現ベクターは、宿主細胞内で複製することもできることが好ましい。発現ベクターは、原核生物であっても真核生物であってもよく、一般にウイルスまたはプラスミドである。発現ベクターには、細菌、真菌、内部寄生虫、節足動物、動物、および植物の細胞を含めた組換え細胞内で機能する(すなわち、直接遺伝子発現)任意のベクターが含まれる。本発明のベクターは、無細胞発現系においてポリペプチドを産生させるために使用することもでき、そのような系は当技術分野で周知である。
【0121】
「作動可能に連結された」は、本明細書で使用する場合、2つ以上の核酸(例えば、DNA)セグメントの間の機能的関係を指す。一般にこれは、転写される配列に対する転写調節エレメントの機能的関係を指す。例えば、プロモーターは、これが、適切な宿主細胞および/または無細胞発現系において、本明細書に定義されるポリヌクレオチドなどのコード配列の転写を刺激または調節する場合、このコード配列に作動可能に連結されている。一般に、転写される配列に作動可能に連結されているプロモーターの転写調節エレメントは、転写される配列に物理的に連続している、すなわち、これらはシス作用である。しかし、エンハンサーなどのいくつかの転写調節エレメントは、これらが転写を増強するコード配列に物理的に連続している、またはごく接近して位置している必要はない。
【0122】
特に、本発明の発現ベクターは、組換え細胞と適合性であり、本発明のポリヌクレオチド分子の発現を制御する制御配列、例えば、転写制御配列、翻訳制御配列、複製起点、および他の制御配列などを含有する。特に、本発明の組換え分子は、転写制御配列を含む。転写制御配列は、転写の開始、伸長、および終止を制御する配列である。特に重要な転写制御配列は、転写開始を制御するもの、例えば、プロモーター、エンハンサー、オペレーター、および抑制因子配列などである。適当な転写制御配列には、本発明の組換え細胞のうちの少なくとも1つにおいて機能することができる任意の転写制御配列が含まれる。様々なそのような転写制御配列は、当業者に既知である。好適な転写制御配列として、細菌、酵母、節足動物、線虫、植物、または動物細胞において機能するもの、例えば、それだけに限らないが、tac、lac、trp、trc、oxy−pro、omp/lpp、rrnB、バクテリオファージλ、バクテリオファージT7、T71ac、バクテリオファージT3、バクテリオファージSP6、バクテリオファージSP01、メタロチオネイン、α−接合因子、ピキア(Pichia)アルコールオキシダーゼ、アルファウイルスサブゲノムプロモーター(シンドビスウイルスサブゲノムプロモーターなど)、抗生物質耐性遺伝子、バキュロウイルス、ヘリオチス・ゼア(Heliothis zea)昆虫ウイルス、ワクシニアウイルス、ヘルペスウイルス、アライグマポックスウイルス、他のポックスウイルス、アデノウイルス、サイトメガロウイルス(中間初期プロモーターなど)、シミアンウイルス40、レトロウイルス、アクチン、レトロウイルスロングターミナルリピート、ラウス肉腫ウイルス、熱ショック、リン酸および硝酸転写制御配列、ならびに原核生物細胞または真核細胞において遺伝子発現を制御することができる他の配列などが挙げられる。
【0123】
宿主細胞
本発明の別の実施形態は、本明細書で記載の1つもしくは複数の組換え分子またはその子孫細胞より形質転換させる宿主細胞を含む。細胞内へのポリヌクレオチド分子の形質転換は、細胞内にポリヌクレオチド分子を挿入することができる任意の方法によって実現することができる。形質転換技法には、それだけに限らないが、トランスフェクション、電気穿孔、微量注入、リポフェクション、吸着、およびプロトプラスト融合が含まれる。組換え細胞は、単細胞のままであることができ、または成長して組織、臓器、もしくは多細胞生物になることができる。形質転換された本発明のポリヌクレオチド分子は、染色体外に残ることができ、または一体化して、発現されるその能力が保持される様式で、形質転換された(すなわち、組換え)細胞の染色体内の1つもしくは複数の部位になることができる。
【0124】
形質転換するのに適した宿主細胞には、本発明のポリヌクレオチドで形質転換され得る任意の細胞が含まれる。本発明の宿主細胞は、本明細書に記載されるポリペプチドを内因的に(すなわち、天然に)産生することができ、または本明細書に記載される少なくとも1つのポリヌクレオチド分子で形質転換された後そのようなポリペプチドを産生することができる。本発明の宿主細胞は、本明細書に定義される少なくとも1つのタンパク質を産生することができる任意の細胞とすることができ、細菌、真菌(酵母を含む)、寄生虫、線虫、節足動物、動物、および植物の細胞を含む。宿主細胞の例として、サルモネラ(Salmonella)、エシェリキア(Escherichia)、バシラス、リステリア(Listeria)、サッカロミセス(Saccharomyces)、スポドプテラ(Spodoptera)、マイコバクテリア(Mycobacteria)、トリコプルシア(Trichoplusia)、BHK(仔ハムスター腎臓)細胞、MDCK細胞、CRFK細胞、CV−1細胞、COS(例えば、COS−7)細胞、およびベロ細胞が挙げられる。宿主細胞のさらなる例は、大腸菌K−12誘導体を含めた大腸菌;チフス菌(Salmonella typhi);弱毒菌株を含めたネズミチフス菌(Salmonella typhimurium);スポドプテラ・フルギペルダ(Spodoptera frugiperda);イラクサギンウワバ(Trichoplusia ni);および非腫瘍形成性マウス筋原細胞G8細胞(例えば、ATCC CRL 1246)である。有用な酵母細胞は、ピシア属(Pichia sp.)、アスペルギルス属(Aspergillus sp.)およびサッカロミセス属(Saccharomyces sp.)を含む。特に好適な宿主細胞は、細菌細胞、酵母細胞または植物細胞である。
【0125】
組換えDNA技術は、例えば、宿主細胞内でのポリヌクレオチド分子のコピー数、これらのポリヌクレオチド分子が転写される効率、得られる転写物が翻訳される効率、および翻訳後修飾の効率を操作することによって、形質転換ポリヌクレオチド分子の発現を改善するのに使用することができる。本発明のポリヌクレオチド分子の発現を増大させるのに有用な組換え技法として、それだけに限らないが、高コピー数プラスミドへのポリヌクレオチド分子の作動可能な連結、1つまたは複数の宿主細胞染色体中へのポリヌクレオチド分子の組込み、プラスミドへのベクター安定性配列の付加、転写制御シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または修飾、翻訳制御シグナル(例えば、リボソーム結合部位、シャイン−ダルガノ配列)の置換または修飾、宿主細胞のコドン使用に対応するための本発明のポリヌクレオチド分子の修飾、および転写物を不安定化する配列の削除が挙げられる。
【0126】
トランスジェニック植物
本発明を実行する際に使用するのに企図されている植物には、単子葉植物および双子葉植物の両方が含まれる。標的植物には、それだけに限らないが、以下のものが含まれる:シリアル(例えば、コムギ、オオムギ、ライムギ、カラスムギ、米、トウモロコシ、ソルガム、および関連作物);ビート(サトウダイコンおよび飼料用ビート);ナシ状果、核果および柔らかい果物(リンゴ、セイヨウナシ、セイヨウスモモ、モモ、アーモンド、サクランボ、イチゴ、ラズベリー、およびブラックベリー);マメ科植物(マメ、レンズマメ、エンドウマメ、ダイズ);油脂植物(ピーナッツ、セイヨウアブラナ、カラシナ、ケシ、オリーブ、ヒマワリ、ココナツ、ヒマシ油植物、カカオ豆、アメリカホドイモ);キュウリ植物(マロー、キュウリ、メロン);繊維植物(綿、アマ、麻、ジュート);柑橘果実(オレンジ、レモン、グレープフルーツ、マンダリン);野菜(ホウレンソウ、レタス、アスパラガス、キャベツ、ニンジン、タマネギ、トマト、ジャガイモ、パプリカ);クスノキ科(lauraceae)(アボカド、シナモン、樟脳);またはタバコ、木の実、コーヒー、サトウキビ、茶、つる植物、ホップ、芝生、バナナ、などの植物、ならびに天然のゴム植物、ならびに観賞植物(花、低木、広葉樹、および針葉樹などの常緑樹)。本発明の標的植物である、アスペルギルス菌種の感染による影響を頻繁に受ける作物には、これに限定されないが、穀類(トウモロコシ、ソルガム、トウジンビエ、イネ、コムギ)、油糧種子(ピーナッツ、ダイズ、ヒマワリ、ワタ)、香辛料(トウガラシ、クロコショウ、コリアンダー、ウコン、ショウガ)および木の実(アーモンド、ピスタチオ、クルミ、ココナツ)が挙げられる。好ましい実施形態では、植物は、製糖用植物、ワタ、トウモロコシ、ソルガム、パイナップル、クリスマスツリーなどの針葉樹、ユーカリノキ、コムギ、カラスムギ、オオムギ、イネおよびキャノーラから選択される。
【0127】
用語「植物」は、本明細書で名詞として使用する場合、植物全体、例えば、商業用の小麦生産用の田畑で成長する植物などを指す。「植物部分」は、植物構造(例えば、葉、幹)、根、花の器官/構造、種子(胚、胚乳、および種皮を含む)、植物組織(例えば、脈管、組織、基本組織など)、細胞、およびこれらの子孫を指す。
【0128】
本発明に照らして定義されるトランスジェニック植物には、所望の植物または植物器官において本発明の少なくとも1つのポリペプチドを産生させるように、組換え技法を使用して遺伝的に改変された植物(ならびに前記植物の部分および細胞)およびその子孫が含まれる。トランスジェニック植物は、A.Slaterら、Plant Biotechnology−The Genetic Manipulation of Plants、Oxford University Press(2003);ならびにP.ChristouおよびH.Klee、Handbook of Plant Biotechnology、John Wiley and Sons(2004)において一般に記載されているものなどの、当技術分野で既知の技法を使用して作製することができる。
【0129】
「トランスジェニック植物」は、同じ種、種類、または栽培品種の野生型植物において見出されない遺伝子コンストラクト(「導入遺伝子」)を含有する。本明細書で呼ばれる「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって生成または変更され、植物細胞内に導入された遺伝子配列を含む。導入遺伝子は、植物細胞に由来する遺伝子配列を含むことができる。一般に、導入遺伝子は、人間の操作、例えば、形質転換などによって植物中に導入されているが、当業者が認識する任意の方法を使用することができる。
【0130】
好ましい実施形態では、トランスジェニック植物は、その子孫が、所望の表現型を分離しないように導入された各遺伝子およびあらゆる遺伝子(導入遺伝子)についてホモ接合である。トランスジェニック植物は、例えば、ハイブリッド種子から成長したF1子孫内などに導入された導入遺伝子(複数可)についてヘテロ接合とすることもできる。そのような植物は、当技術分野で周知であるハイブリッドの活力(hybrid vigour)などの利点をもたらすことができる。
【0131】
本発明のポリヌクレオチドは、発生のすべての段階の間に、トランスジェニック植物内で構成的に発現され得る。植物または植物器官の使用に応じて、ポリペプチドは、段階特異的な様式で発現され得る。さらに、ポリヌクレオチドは、組織特異的に発現され得る。
【0132】
植物内で対象とするポリペプチドをコードする遺伝子の発現を引き起こすことが知られているか、または見出されている制御配列を、本発明において使用することができる。使用される制御配列の選択は、対象とする標的植物および/または標的器官に依存する。そのような制御配列は、植物または植物ウイルスから得ることができ、または化学的に合成することができる。そのような制御配列は、当業者に周知である。
【0133】
植物細胞の安定なトランスフェクション、またはトランスジェニック植物の確立に適したいくつかのベクターは、例えば、Pouwelsら、Cloning Vectors:A Laboratory Manual(1985、補遺1987);WeissbachおよびWeissbach、Methods for Plant Molecular Biology、Academic Press(1989);ならびにGelvinら、Plant Molecular Biology Manual、Kluwer Academic Publishers(1990)において記載されている。一般に、植物発現ベクターは、5’および3’制御配列の転写制御下の1つまたは複数のクローン化植物遺伝子、および優性選択マーカーを含む。そのような植物発現ベクターは、プロモーター調節領域(例えば、誘導性もしくは構成的な、環境的もしくは発生的に制御された、または細胞もしくは組織特異的発現を制御する調節領域)、転写開始部位、リボソーム結合部位、RNA処理シグナル、転写終止部位、および/またはポリアデニル化シグナルも含有することができる。
【0134】
植物細胞内で活性であるいくつかの構成的プロモーターが記載されている。植物内の構成的発現に適したプロモーターとして、それだけに限らないが、カリフラワーモザイクウイルス(CaMV)35Sプロモーター、ゴマノハグサモザイクウイルス(FMV)35S、サトウキビ桿菌状ウイルスプロモーター、ツユクサ(commelina)黄斑ウイルスプロモーター、リブロース−1,5−ビス−リン酸カルボキシラーゼの小サブユニットに由来する光誘導プロモーター、イネサイトゾルトリオースリン酸イソメラーゼプロモーター、シロイヌナズナ(Arabidopsis)のアデニンホスホリボシルトランスフェラーゼプロモーター、イネアクチン1遺伝子プロモーター、マンノピン合成酵素およびオクトピン合成酵素プロモーター、Adhプロモーター、スクロース合成酵素プロモーター、R遺伝子複合体プロモーター、およびクロロフィルα/β結合タンパク質遺伝子プロモーターが挙げられる。これらのプロモーターは、植物内で発現されたDNAベクターを作り出すのに使用されている;例えば、WO84/02913を参照。これらのプロモーターのすべては、様々な型の植物で発現可能な組換えDNAベクターを作り出すのに使用されている。
【0135】
植物の起源組織、例えば、葉、種子、根、または茎などにおける発現の目的のために、本発明において利用されるプロモーターは、これらの特定の組織において相対的に高い発現を有することが好適である。この目的のために、組織もしくは細胞特異的または組織もしくは細胞で増強される発現を伴う遺伝子についてのいくつかのプロモーターから1つを選択することができる。文献で報告されたそのようなプロモーターの例として、エンドウマメに由来する葉緑体グルタミン合成酵素GS2プロモーター、コムギに由来する葉緑体フルクトース−1,6−ビホスファターゼ(biphosphatase)プロモーター、ジャガイモに由来する核光合成ST−LS1プロモーター、シロイヌナズナ(Arabidopsis thaliana)に由来するセリン/スレオニンキナーゼプロモーター、およびグルコアミラーゼ(CHS)プロモーターが挙げられる。イースタンラーチ(アメリカカラマツ(Larix laricina))に由来するリブロース−1,5−二リン酸カルボキシラーゼプロモーター、マツに由来するCab遺伝子、Cab6についてのプロモーター、コムギに由来するCab−1遺伝子についてのプロモーター、ホウレンソウに由来するCab−1遺伝子についてのプロモーター、イネに由来するCab1R遺伝子についてのプロモーター、トウモロコシ(Zea mays)に由来するピルビン酸オルトリン酸ジキナーゼ(PPDK)プロモーター、タバコLhcb1*2遺伝子についてのプロモーター、シロイヌナズナSuc2スクロース−H30共輸送体プロモーター、ホウレンソウに由来するチラコイド膜タンパク質についてのプロモーター(PsaD、PsaF、PsaE、PC、FNR、AtpC、AtpD、Cab、RbcS)も光合成的に活性な組織において活性であると報告されている。
【0136】
クロロフィルα/β結合タンパク質についての他のプロモーター、例えば、白ガラシ(シナピス・アルバ(Sinapis alba))に由来するLhcB遺伝子およびPsbP遺伝子についてのプロモーターなども本発明において利用することができる。(1)熱、(2)光(例えば、エンドウマメRbcS−3Aプロモーター、トウモロコシRbcSプロモーター);(3)アブシシン酸などのホルモン(4)創傷(例えば、WunI);もしくは(5)化学物質、例えば、メチルジャスミネート(jasminate)、サリチル酸、ステロイドホルモン、アルコール、セーフナー(WO97/06269)などによって調節されるプロモーターを含めた、環境、ホルモン、化学および/または発生シグナルに対する応答において調節される様々な植物遺伝子プロモーターも植物細胞内のRNA結合タンパク質遺伝子の発現のために使用することができ、または(6)器官特異的なプロモーターを使用することも有利となり得る。
【0137】
植物のシンク組織、例えば、ジャガイモ植物の塊茎、トマトの果実、またはダイズ、アブラナ、綿、トウモロコシ、コムギ、イネ、およびオオムギの種子などにおける発現の目的のために、本発明において利用されるプロモーターは、これらの特定の組織において相対的に高い発現を有することが好適である。クラスIパタチンプロモーター、大小サブユニットの両方のジャガイモ塊茎ADPGPP遺伝子についてのプロモーター、スクロース合成酵素プロモーター、22kDのタンパク質複合体およびプロテイナーゼ阻害剤を含めた主要な塊茎タンパク質についてのプロモーター、顆粒結合型デンプン合成酵素遺伝子(GBSS)についてのプロモーター、ならびに他のクラスIおよびIIパタチンプロモーターを含めて、塊茎特異的または塊茎で増強される発現を伴う遺伝子についてのいくつかのプロモーターが知られている。他のプロモーターも、種子または果実などの特定の組織内でタンパク質を発現させるのに使用することができる。β−コングリシニンについてのプロモーター、または他の種子特異的なプロモーター、例えば、ナピンおよびファセオリンプロモーターなどを使用することができる。トウモロコシ胚乳発現について特に好適なプロモーターは、イネに由来するグルテリン遺伝子についてのプロモーター、より具体的には、Osgt−1プロモーターである。コムギにおける発現に適したプロモーターの例には、ADPグルコースピロシンターゼ(pyrosynthase)(ADPGPP)サブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素および脱分枝酵素、胚形成主要タンパク質、グリアジン、ならびにグルテニンについてのプロモーターが含まれる。イネにおけるかかるプロモーターの例には、ADPGPPサブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素、脱分岐酵素、スクロース合成酵素ならびにグルテリンについてのプロモーターが挙げられる。特に好ましいプロモーターは、イネグルテリンについてのプロモーター、すなわちOsgt−1遺伝子である。オオムギについてのかかるプロモーターの例には、ADPGPPサブユニット、顆粒結合型および他のデンプン合成酵素、分岐酵素、脱分岐酵素、スクロース合成酵素、ホルデイン、胚グロブリンならびにアリューロン特異的なタンパク質についてのプロモーターが挙げられる。
【0138】
根に特異的なプロモーターも使用することができる。そのようなプロモーターの例は、酸性キチナーゼ遺伝子についてのプロモーターである。根組織における発現は、同定されたCaMV35Sプロモーターの根に特異的なサブドメインを利用することによっても実現することができる。
【0139】
5’非翻訳リーダー配列は、本発明のポリヌクレオチドの異種遺伝子配列を発現するように選択されたプロモーターから得ることができ、必要に応じてmRNAの翻訳を増大するように特異的に改変することができる。導入遺伝子の発現を最適化することの概説については、Kozielら(1996)を参照。5’非翻訳領域は、適当な真核生物遺伝子、植物遺伝子(コムギおよびトウモロコシクロロフィルa/b結合タンパク質遺伝子リーダー)に由来する植物ウイルスRNA(とりわけ、タバコモザイクウイルス、タバコエッチウイルス、トウモロコシ萎縮モザイクウイルス、アルファルファモザイクウイルス)、または合成遺伝子配列から得ることもできる。本発明は、非翻訳領域が、プロモーター配列を伴う5’非翻訳配列に由来するコンストラクトに限定されない。リーダー配列は、無関係のプロモーターまたはコード配列に由来する場合もある。本発明の脈絡において有用なリーダー配列は、トウモロコシHsp70リーダー(US5,362,865およびUS5,859,347)、およびTMVΩ要素を含む。
【0140】
転写の終止は、対象とするポリヌクレオチドにキメラベクター中で作動可能に連結された3’非翻訳DNA配列によって実現される。組換えDNA分子の3’非翻訳領域は、RNAの3’末端にアデニル酸ヌクレオチドの付加を引き起こすように植物内で機能するポリアデニル化シグナルを含有する。3’非翻訳領域は、植物細胞内で発現される様々な遺伝子から得ることができる。ノパリン合成酵素3’非翻訳領域、エンドウマメ小サブユニットRubisco遺伝子に由来する3’非翻訳領域、ダイズ7S種子貯蔵タンパク質遺伝子に由来する3’非翻訳領域は一般に、この能力において使用される。アグロバクテリウム(Agrobacterium)腫瘍誘導(Ti)プラスミド遺伝子のポリアデニル酸シグナルを含有する3’転写非翻訳領域も適している。
【0141】
細胞内に遺伝子を直接送達するための4つの一般的な方法、すなわち、(1)化学的方法(Grahamら、1973);(2)微量注入などの物理的方法(Capecchi、1980);電気穿孔(例えば、WO87/06614、US5,472,869、5,384,253、WO92/09696、およびWO93/21335を参照);および遺伝子銃(例えば、US4,945,050およびUS5,141,131を参照);(3)ウイルスベクター(Clapp、1993;Luら、1993;Eglitisら、1988);ならびに(4)受容体媒介機構(Curielら、1992;Wagnerら、1992)が記載されている。
【0142】
使用することができる加速方法には、例えば、微粒子銃などが含まれる。植物細胞に形質転換核酸分子を送達するための方法の一例は、微粒子銃である。この方法は、Yangら、Particle Bombardment Technology for Gene Transfer、Oxford Press、Oxford、England(1994)によって概説されている。非生物学的粒子(マイクロプロジェクタイル)は、核酸をコーティングし、推進力によって細胞内に送達する。例示的な粒子には、タングステン、金、白金などからなるものが含まれる。微粒子銃が単子葉植物を再現性よく形質転換する有効な手段であることに加えて、微粒子銃の特定の利点は、プロトプラストの分離もアグロバクテリウム感染の感受性も必要とされないことである。加速によってトウモロコシ細胞内にDNAを送達するための方法の例示的な実施形態は、微粒子銃α粒子送達システムであり、これは、DNAでコーティングされた粒子を、ステンレス鋼またはNytexスクリーンなどのスクリーンを通して、懸濁液中で培養されたトウモロコシ細胞で覆われたフィルター表面上に推進するのに使用することができる。本発明とともに使用するのに適した粒子送達システムは、ヘリウム加速PDS−1000/He銃であり、Bio−Rad Laboratoriesから入手可能である。
【0143】
照射のために、懸濁液中の細胞をフィルター上で濃縮することができる。照射される細胞を含有するフィルターは、マイクロプロジェクタイル停止プレートの下に適切な距離をおいて配置される。必要に応じて、1つまたは複数のスクリーンも、銃と照射される細胞との間に配置される。
【0144】
あるいは、未成熟胚または他の標的細胞を、固体培地上に配置することができる。照射される細胞は、マイクロプロジェクタイル停止プレートの下に適切な距離をおいて配置される。必要に応じて、1つまたは複数のスクリーンも、加速デバイスと照射される細胞との間に配置される。本明細書に示した方法を使用することによって、1000またはそれ以上の焦点の、マーカー遺伝子を一過性に発現する細胞を得ることができる。照射して48時間後に外来遺伝子産物を発現する焦点内の細胞数は、1〜10個、平均で1〜3個の範囲であることが多い。
【0145】
照射形質転換において、照射前の培養条件および照射パラメータを最適化することによって、最大数の安定な形質転換体を得ることができる。照射についての物理的および生物学的パラメータの両方が、この技術において重要である。物理的要因は、DNA/マイクロプロジェクタイル沈殿物の操作を伴うもの、またはマクロもしくはマイクロプロジェクタイルの飛行および速度に影響するものである。生物学的要因には、照射前および照射直後の細胞の操作に関与するすべてのステップ、照射に伴う外傷の軽減に役立てるための標的細胞の浸透圧調整、ならびに直線化DNAまたは無傷のスーパーコイルプラスミドなどの形質転換DNAの性質が含まれる。照射前の操作は、未成熟胚の形質転換の成功に特に重要であると考えられている。
【0146】
別の代替の実施形態では、プラスチドを安定に形質転換することができる。高等植物におけるプラスチド形質転換について開示された方法には、選択マーカーを含有するDNAの粒子銃送達、および相同的組換えを通じたプラスチドゲノムへのDNAの標的化が含まれる(US5,451,513、US5,545,818、US5,877,402、US5,932479、およびWO99/05265)。
【0147】
したがって、条件を完全に最適化するための小規模研究において、照射パラメータの様々な態様を調整することが望まれる場合があることが企図されている。物理的パラメータ、例えば、ギャップ距離、飛行距離、組織距離、およびヘリウム圧力などを調整することが特に望まれる場合がある。レシピエント細胞の生理的状態に影響し、したがって形質転換および組込み効率に影響し得る条件を修正することによって外傷悪化要因を最小限にすることもできる。例えば、レシピエント細胞の浸透圧状態、組織水和、および継代培養段階または細胞周期を、最適な形質転換のために調整することができる。他の通常の調整を実施することも、本開示を踏まえると当業者に分かるであろう。
【0148】
アグロバクテリウム媒介トランスファーは、植物細胞内に遺伝子を導入するために広く適用可能なシステムであり、その理由は、DNAを全植物組織内に導入し、それによってプロトプラストから無傷の植物を再生する必要を回避することができるためである。植物細胞内にDNAを導入するために、ベクターを組み込んでいるアグロバクテリウム媒介植物を使用することは、当技術分野で周知である(例えば、US5,177,010、US5,104,310、US5,004,863、US5,159,135を参照)。さらに、T−DNAの組込みは、比較的正確なプロセスであり、ほとんど再配列をもたらさない。トランスファーされるDNAの領域は、ボーダー配列によって画定され、介在DNAは、植物ゲノム内に通常挿入される。
【0149】
最新のアグロバクテリウム形質転換ベクターは、(Kleeら、Plant DNA Infectious Agents、HohnおよびSchell編、Springer−Verlag、New York、179〜203頁(1985))に記載されているように、大腸菌ならびにアグロバクテリウム内で複製することができ、好都合な操作を可能にしている。さらに、アグロバクテリウム媒介遺伝子トランスファーのためのベクターにおける技術的進歩は、ベクター中の遺伝子および制御部位の配置を改善することによって、様々なポリペプチドをコードする遺伝子を発現することができるベクターの構築を促進している。記載されたベクターは、挿入されたポリペプチドコード遺伝子を直接発現させるために、プロモーターおよびポリアデニル化部位に隣接して、好都合なマルチリンカー領域を有し、本目的に適している。さらに、アームドおよびディスアームドTi遺伝子(armed and disarmed Ti gene)を含有するアグロバクテリウムを、形質転換のために使用することができる。アグロバクテリウム媒介形質転換が効率的である植物の種類において、遺伝子トランスファーの容易で明確な性質のために、これが最適の方法である。
【0150】
アグロバクテリウム形質転換法を使用して形成されたトランスジェニック植物は一般に、1つの染色体上に1つの遺伝子座を含有する。そのようなトランスジェニック植物は、付加された遺伝子についてヘミ接合であると呼ぶことができる。より好適なのは、付加された構造遺伝子についてホモ接合であるトランスジェニック植物、すなわち、染色体対の各染色体上の同じ座位に1つの遺伝子という、2つの付加された遺伝子を含有するトランスジェニック植物である。ホモ接合トランスジェニック植物は、1つの付加された遺伝子を含有する独立した分離個体のトランスジェニック植物を有性交配(自家受粉)し、産生された種子のいくつかを発芽させ、対象とする遺伝子について得られた植物を分析することによって得ることができる。
【0151】
2つの異なるトランスジェニック植物を交配することによって、2つの独立して分離している外来遺伝子を含有する子孫を産生させることもできることも理解されるべきである。適切な子孫を自家受粉することにより、両方の外来遺伝子についてホモ接合である植物を産生させることができる。親植物への戻し交配、および非トランスジェニック植物との外交配も、栄養繁殖のように企図されている。様々な形質および作物に一般に使用される他の育種法の記述は、Fehr、Breeding Methods for Cultivar Development、Wilcox J.編、American Society of Agronomy、Madison Wis(1987)に見出すことができる。
【0152】
植物プロトプラストの形質転換は、リン酸カルシウム沈殿、ポリエチレングリコール処理、電気穿孔、およびこれらの処理の組合せに基づく方法を使用して実現することができる。様々な植物の種類にこれらのシステムを適用することは、プロトプラストからその特定の植物系統を再生する能力に依存する。プロトプラストから穀類を再生するための例示的方法が記載されている(Fujimuraら、1985;Toriyamaら、1986;Abdullahら、1986)。
【0153】
細胞形質転換の他の方法も使用することができ、これには、それだけに限らないが、花粉中にDNAを直接トランスファーすること、植物の生殖器中にDNAを直接注射すること、または未成熟胚の細胞内にDNAを直接注射し、その後乾燥された胚を再水和することによる植物中へのDNAの導入が含まれる。
【0154】
1つの植物プロトプラスト形質転換体、または様々な形質転換された外植片から植物を再生し、発生させ、培養することは、当技術分野で周知である(Weissbachら、Methods for Plant Molecular Biology、Academic Press、San Diego、Calif.(1988))。この再生および成長プロセスは一般に、形質転換細胞を選択するステップ、胚発生の通常の段階を介し、根付いた小植物段階を介して個別化した細胞を培養するステップを含む。トランスジェニック胚および種子も同様に再生される。得られたトランスジェニックの根付いた苗条は、土壌などの適切な植物成長培地にその後植えられる。
【0155】
異種の、外来遺伝子を含有する植物の発生または再生は、当技術分野で周知である。再生された植物は、自家受粉することによって、ホモ接合トランスジェニック植物をもたらすことが好ましい。さもなければ、再生された植物から得られる花粉は、農学的に重要な系統の種子から成長した植物と交雑される。反対に、これらの重要な系統の植物からの花粉は、再生された植物に授粉するのに使用される。所望の外因性核酸を含有する本発明のトランスジェニック植物は、当業者に周知の方法を使用して栽培される。
【0156】
主にアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を使用することによって双子葉植物を形質転換し、トランスジェニック植物を得るための方法は、綿(US5,004,863、US5,159,135、US5,518,908);ダイズ(US5,569,834、US5,416,011);アブラナ属(Brassica)(US5,463,174);ピーナッツ(Chengら、1996);およびエンドウマメ(Grantら、1995)について公開されている。
【0157】
外因性核酸を導入することによって植物中に遺伝的変異を導入するため、およびプロトプラストまたは未成熟植物胚から植物を再生するために、コムギおよびオオムギなどの穀類植物を形質転換するための方法は、当技術分野で周知である。例えば、CA2,092,588、AU61781/94、AU667939、US6,100,447、WO97/048814、US5,589,617、US6,541,257およびWO99/14314を参照。トランスジェニックコムギまたはオオムギ植物は、アグロバクテリウム・ツメファシエンス媒介形質転換手順によって産生されることが好ましい。所望の核酸コンストラクトを担持するベクターは、組織培養された植物もしくは外植片の再生可能なコムギ細胞、またはプロトプラストなどの適当な植物系内に導入することができる。
【0158】
再生可能なコムギ細胞は好ましくは、未成熟胚、成熟胚の胚盤、これらの組織または分裂組織から得られるカルスに由来する。
【0159】
トランスジェニック細胞および植物内に導入遺伝子が存在することを確認するために、ポリメラーゼ連鎖反応(PCR)増幅またはサザンブロット分析を、当業者に既知の方法を使用して実施することができる。導入遺伝子の発現産物は、産物の性質に応じて様々な方法のうちのいずれでも検出することができ、ウエスタンブロットおよび酵素アッセイを含む。タンパク質発現を定量化し、様々な植物組織における複製を検出するための1つの特に有用な方法は、GUSなどのレポーター遺伝子を使用することである。トランスジェニック植物が得られたら、これらを成長させることによって、所望の表現型を有する植物組織または植物部分を生成することができる。植物組織または植物部分を回収することができ、かつ/または種子を収集することができる。種子は、所望の特徴を有する組織または部分を伴った追加の植物を成長させるための供給源として機能を果たすことができる。
【0160】
ある実施形態では、本発明のトランスジェニック植物は、US6,369,299に一般的に記載された方法を使用して作製される。
【0161】
トランスジェニック非ヒト動物
「トランスジェニック非ヒト動物」は、同じ種または品種の野生型動物において見出されない遺伝子コンストラクト(「導入遺伝子」)を含有する、ヒト以外の動物を指す。本明細書で呼ばれる「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって生成または変更され、動物細胞内に導入された遺伝子配列を含む。導入遺伝子は、動物細胞に由来する遺伝子配列を含むことができる。一般に、導入遺伝子は、人間の操作、例えば、形質転換などによって動物中に導入されているが、当業者が認識する任意の方法を使用することができる。
【0162】
トランスジェニック動物を作製するための技法は、当技術分野で周知である。この課題に対する有用な一般的な教科書は、Houdebine、Transgenic animals−Generation and Use(Harwood Academic、1997)である。
【0163】
異種のDNAを、例えば、哺乳動物の受精卵子中に導入することができる。例えば、全能性または多能性幹細胞を、微量注入、リン酸カルシウム媒介沈殿、リポソーム融合、レトロウイルス感染または他の手段によって形質転換することができる。次いで形質転換細胞は、胚内に導入され、次いで胚は、発達してトランスジェニック動物になる。非常に好適な方法では、発達中の胚に、所望のDNAを含有するレトロウイルスを感染させ、感染した胚からトランスジェニック動物が生じる。しかし、もっとも好適な方法では、適切なDNAが、好ましくは単細胞段階で胚の前核または細胞質中に同時注入され、胚を発達させて成熟したトランスジェニック動物にする。
【0164】
トランスジェニック動物を作製するのに使用される別の方法は、標準的な方法によって前核段階の卵子中に核酸を微量注入することを伴う。次いで注入された卵子は、培養された後、偽妊娠のレシピエントの輸卵管に移される。
【0165】
トランスジェニック動物は、核トランスファー技術によっても作製することができる。この方法を使用して、ドナー動物に由来する線維芽細胞は、制御配列の制御下で、対象とする結合ドメインまたは結合パートナーについてのコード配列を組み込んでいるプラスミドで安定にトランスフェクトされる。次いで安定なトランスフェクタントは、除核された卵母細胞に融合され、培養され、メスのレシピエントに移される。
【0166】
組成物
本発明の組成物は、「許容可能な担体」とも本明細書で呼ばれる、賦形剤を含む。賦形剤は、処置される動物、植物、植物もしくは動物物質、または環境(土壌および水試料を含む)が耐容性を示すことができる任意の物質とすることができる。そのような賦形剤の例として、水、食塩水、リンガー液、デキストロース溶液、ハンクス液、および他の水性生理学的平衡塩溶液が挙げられる。非水ビヒクル、例えば、固定油、ゴマ油、オレイン酸エチル、または中性脂肪なども使用することができる。他の有用な製剤は、粘度増強剤、例えば、ナトリウムカルボキシメチルセルロース、ソルビトール、またはデキストランなどを含有する懸濁液を含む。賦形剤は、等張性および化学的安定性を増強する物質などの軽微な量の添加剤も含有することができる。緩衝液の例には、リン酸緩衝液、炭酸水素塩緩衝液、およびTris緩衝液が含まれ、一方、保存剤の例には、チメロサール、またはo−クレゾール、ホルマリン、およびベンジルアルコールが含まれる。賦形剤は、組成物、例えば、それだけに限らないが、ポリマー制御放出ビヒクル、生分解性インプラント、リポソーム、細菌、ウイルス、他の細胞、油、エステル、およびグリコールの半減期を増大させるのにも使用することができる。
【0167】
ある実施形態では、本発明のポリペプチドは、固体支持体上に固定される。これにより、s−トリアジンまたはジアジンの加水分解速度および/または程度を増強でき、および/またはポリペプチドの安定性を高めることができる。例えば、ポリペプチドは、ポリウレタン基質上に固定化し(Gordonら、1999)、または適切なリポソーム中にカプセル化することができる(Petrikovicsら、2000aおよびb)。ポリペプチドは、消火において日常的に使用されるものなどのフォームを含む組成物中に組み込むこともできる(LeJeuneら、1998)。当業者に理解されるように、本発明のポリペプチドは、WO00/64539に開示のようにスポンジまたはフォームですぐに使用することができる。本発明に有用な他の固体支持体には、アクリル型構造を有する樹脂、セパビーズEC−EP(Resindion srl−−三菱化学株式会社)およびEupergit C(Rohm−Degussa)などのエポキシ官能基を有する樹脂、またはセパビーズEC−hasおよびEC−EA(Resindion srl−−三菱化学株式会社)などの第1級アミノ基を有する樹脂が挙げられる。いずれの場合も、ポリペプチドは、樹脂と接触させられ、酵素がマトリックスに結合するように、官能基(エポキシド)の高い反応性、またはグルタルアルデヒドなどの二官能性作用剤による樹脂の活性化により固定される。本発明に適した他の樹脂は、ポリスチレン樹脂、巨大網状樹脂およびセパビーズEC−Q1Aなどの塩基性官能基を有する樹脂であり、ポリペプチドは樹脂上に吸収され、次いで二官能性作用剤(グルタルアルデヒド)との架橋結合によって安定化する。
【0168】
ある実施形態では、組成物は、Zn2+および/またはCo2+を含む。別の実施形態では、s−トリアジンまたはジアジンを加水分解するための本発明の方法は、本発明のポリペプチドについての補助因子としてZn2+および/またはCo2+を提供するステップを含む。
【0169】
本発明の一実施形態は、本発明の組成物を、動物、植物、動物もしくは植物物質、または環境(土壌および水試料を含む)中に徐々に放出することができる制御放出製剤である。本明細書で使用する場合、制御放出製剤は、制御放出ビヒクル中に本発明の組成物を含む。適当な制御放出ビヒクルとして、それだけに限らないが、生体適合性ポリマー、他のポリマー基質、カプセル、マイクロカプセル、微粒子、大量瞬時投与製剤、浸透圧ポンプ、拡散デバイス、リポソーム、リポスフェア、および経皮送達システムが挙げられる。好適な制御放出製剤は、生分解性(すなわち、生体内分解性)である。
【0170】
本発明の好適な制御放出製剤は、S−トリアジンまたはジアジンを含む範囲内にある土壌または水中に本発明の組成物を放出することができる。この製剤は好ましくは、約1〜約12カ月の範囲の時間にわたって放出される。本発明の好適な制御放出製剤は、好ましくは少なくとも約1カ月間、より好ましくは少なくとも約3カ月間、さらにより好ましくは少なくとも約6カ月間、さらにより好ましくは少なくとも約9カ月間、さらにより好ましくは少なくとも約12カ月間処理を行うことができる。
【0171】
S−トリアジンまたはジアジンを加水分解するための有効な組成物を生成するのに必要とされる本発明のポリペプチド、ベクター、または宿主細胞などの濃度は、除染される試料の性質、試料中のS−トリアジンまたはジアジンの濃度、ならびに組成物の製剤に依存する。組成物内のポリペプチド、ベクター、または宿主細胞などの有効濃度は、本発明の方法を用いて実験的に容易に求めることができる。
【0172】
本発明の酵素および/またはそれをコードする宿主細胞は、WO2004/112482およびWO2005/26269に一般に記載されているように、コーティング組成物で使用することができる。
【実施例1】
【0173】
増強した活性を有するTrzN突然変異体
方法および材料
トランケートしたpET14bプラスミド(pETcc2)を、TrzNおよびその変異体の発現のために使用した。ユニークなNdeIおよびBamHI部位を用いて、全trzN遺伝子をpETcc2中にクローニングし、大腸菌株BL21λDE3(Novagen)中で発現させた。NdeI/BamHI消化型pETcc2をpETcc2::egfp(図2)から調製し、図2ではライブラリーにおける再連結したベクター(すなわち、青色光下で強く蛍光を発する連結したベクター)の比率の単純なビジュアル表示を提供した。
【0174】
コドンを最適化したTrzN(配列番号2−TrzNco)を、GENEART AG(BioPark、Josef−Engert−Str.11、D−93053 レーゲンブルグ、ドイツ)によって産生した。
【0175】
ランダムな変異誘発を、製品説明書に従い、GeneMorphII(Stratagen)を用いて行った。オリゴヌクレオチドプライマー1および2を、遺伝子の増幅に使用した(表2)。
【0176】
表2に詳述のように、部位飽和変異誘発(Site-saturation mutagenesis)を、プライマー3〜24を用いて、PCR媒介型部位特異的変異誘発によって行った。NNS縮重(Georgescuら、Directed Evolution Library Creation、編:F.H.Arnold、G.Georgiou、Humana Press、Totowa、NJ、2003、pp.75〜89)を、trzN遺伝子の67番目、91番目、131番目、159番目、161番目、243番目、246番目、294番目、335番目および350番目コドンで多様性を生じさせるために使用した(ここでは、Nは任意のヌクレオチドであり、SはGまたはCのいずれかである)。
【0177】
ランダムな部位飽和したライブラリーを、アンピシリン200μg.mL−1、IPTG1μMを補充したLB寒天プレートを用いて選別し、アトラジン1mg.mL−1(90%アトラジンw/w;Gesaprim 900 WG、Syngenta)を浸透させた。TrzN活性を、アトラジン脱塩素化によって評価し、結果として活性TrzNを発現するコロニー付近の培地の清澄化が生じた。活性レベルを、清澄化が起きる速度によって決定した。
【0178】
全配列決定を、Micromon(モナッシュ大学、メルボルン、ビクトリア)によって行った。
【0179】
以前に報告されたように(Shapirら、2005b)、264nMでの吸光度の減少により測定するとき、His6TrzNcc3.2を精製することにより、基質の加水分解速度を測定することによって、基質の範囲を試験した。アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンおよびシアノジン(>99%純度Pestinal標準;Sigma)を、この方法を用いて試験した。His6TrzNcc3.2をアフィニティークロマトグラフィー(HisTrap;GE HealthCare)、続いてサイズ排除クロマトグラフィー(Superdex200;GE HealthCare)を用いて精製した。
【0180】
【表2】
【0181】
結果
反復性ランダム変異誘発
アトラジンのプレート清澄化アッセイを、アトラジンを加水分解的に脱塩素化するBL21λDE3pETcc2::trzNcoの能力を評価するために使用した。30日後に、細菌増殖付近における固体培地の清澄化は観察されなかったため、BL21λDE3pETcc2::trzNcoは、アトラジンのクロロヒドロラーゼ活性レベルは、無かったか検出できなかったと考えられた。trzNcoのランダムな突然変異体を、ローフィデリティのDNAポリメラーゼMutazyme II(GeneMorph II)を用いて作製し、アトラジンに対する加水分解活性について、寒天プレートを含むアトラジンにおいて選別した。trzNcoにおける単一の同義突然変異(T468C)により、156番目のコドンをコードするフェニルアラニンがTTTからTTCに変わった(配列番号3)。突然変異体(TrzN L1、表3)は、37℃で8日後にアトラジンを清澄化する能力がもたらされた。
【0182】
ランダムな変異誘発の次のラウンドについて、TrzN L1を鋳型として使用した(反復1)。27個の突然変異体において、親trzNcoより素早く清澄ゾーンを形成する能力がBL21にもたらされることを発見した(TrzN L1、表3)。清澄ゾーンは、反復1の突然変異体について、3から6日の間で出現した。清澄ゾーンをもっとも素早く形成する15個(TrzN cc1.1、TrzN cc1.3、TrzN cc1.4、TrzN cc1.6、TrzN cc1.8、TrzN cc1.9、TrzN cc1.10、TrzN cc1.11、TrzN cc1.12、TrzN cc1.13、TrzN cc1.14、TrzN cc1.15、TrzN cc1.25、TrzN cc1.26およびTrzN cc1.27)の突然変異体を、ランダム変異誘発の別のラウンドのための鋳型として使用した(反復2)。
【0183】
全体で最良に実行できる突然変異体を選択できるように、反復2の変異誘発PCRからの産物を一緒に選別した。45〜96時間で清澄ゾーンを形成する36個のコロニーを選択した(表3)。反復2の最良の13個の突然変異体(45〜48時間の間で清澄化する;TrzN cc2.1、TrzN cc2.2、TrzN cc2.3、TrzN cc2.4、TrzN cc2.5、TrzN cc2.6、TrzN cc2.7、TrzN cc2.8、TrzN cc2.9、TrzN cc2.10、TrzN cc2.11、TrzN cc2.12およびTrzN cc2.13)を、ランダム変異誘発の3回目の反復のための鋳型として使用した。
【0184】
【表3−1】
【表3−2】
【0185】
再び、全体で最良に実行できる突然変異体を選択できるように、反復2の変異誘発PCRからの産物を一緒に選別した。3回目の反復の13個の突然変異体を選択し、24〜31時間の清澄化時間をもたらすものを選択した。突然変異体におけるヌクレオチドおよびアミノ酸の置換は、表4および5にそれぞれまとめる。単一遺伝子内でもっとも多いヌクレオチド突然変異は、14個であり、任意の変異体酵素において最大7つのアミノ酸置換であった。
【0186】
【表4】
【0187】
【表5−1】
【表5−2】
【0188】
興味深いことに、両方の突然変異が、アミノ酸置換をもたらし、清澄化速度に影響を与えないようであったが、これは、タンパク質のフォールディングおよび安定性に影響する因子に加えて、全体のTrzN活性の転写または翻訳の決定因子が存在することを示唆する。このことは、タンパク質配列は変化させなかったが、むしろF156をコードするのに使用したフェニルアラニンのコドンを(TTTからTTCに)変えた、trzN L1突然変異(T468C)へのtrzNcoの作用によって、明らかに実際に示される。それらが同一産物であるにも関わらず、trzN L1は、明らかにその親遺伝子より優れていた(表3)。
【0189】
部位飽和変異誘発
ランダム変異誘発プログラムの1回目、2回目もしくは3回目の反復の間で、1回より多く変化した、またはランダム変異誘発の2回目および3回目の反復に強く選択された、またはランダム変異誘発の1つのラウンドに存在し、独立した2回目の部位突然変異として、ランダム変異誘発の2回目および3回目の反復の親鋳型において他の突然変異と組み合わせたアミノ酸は、trzNco突然変異体の改良したアトラジンデクロリナーゼ活性の重要な決定因子であると考えられた。ランダム変異誘発によって導入されそうにないアミノ酸との置換を含む、これらの位置の配列スペースをより完全に探索するために、部位飽和ライブラリーをこれらの各部位(コドン67、91、131、159、161、210、243、246、294、335および350;表6)について作成した。
【0190】
【表6】
【0191】
いくつかの部位での別のアミノ酸の広いスペクトルによるアミノ酸置換により、親遺伝子と比較して、清澄化速度が上昇する結果となった一方、他のものは、1つの別のアミノ酸のみへのかなり保存的な変化を許すのみであった。12個(P、N、T、D、V、G、C、S、Q、H、YまたはI)、6個(D、S、E、K、VまたはA)および5個(Y、N、F、KまたはH)の別のアミノ酸に対するL131、D350およびG246のそれぞれの置換により、野生型酵素と比較して増加したデクロリナーゼ活性がもたらされた。A294は、3つの別のアミノ酸(T、SまたはL)によって置換し、より高いアトラジンデクロリナーゼ活性を産生することができた一方、A159およびL243は、2つの別の各アミノ酸(それぞれVまたはT、およびPまたはG)のみによってうまく置換することができた。5つの位置で、1つの可能性のある置換のみが、改良された活性を産生した(Y67F、T91S、S161G、P210AおよびL335M)(表6)。我々はまた、L131P A159VバックグラウンドにおけるD38X部位飽和ライブラリーから100個のコロニーを選別した。我々は、明らかに改良された清澄化の表現型を有する5個のコロニーを得た。かかる表現型をもたらす置換は、D38Nのみであった。
【0192】
変異体Trz Ncc3.2における突然変異の特性評価
SDS−PAGEおよび動力学的解析により、かなり大部分の活性の改良が、増加した発現の結果であることが実際に示された。実際に、発現は、変異誘発の各ラウンドで徐々に増加し、3回目のラウンドで得たもっとも活性の高い突然変異体は、3つのアミノ酸置換(D38N、 L131PおよびA159V)を含んだ(図3)。trzNにおいて多くのサイレント突然変異があったが、これらは、tRNAの利用、mRNAの安定性または翻訳効率への二次構造の影響の変化により、発現の増加に寄与したようである。各アミノ酸変化の溶解性の増加への寄与は、これらをTrzN野生型バックグラウンドに個々に導入することにより、および逆方向突然変異を突然変異体に個々に導入することにより評価した。A159V置換の存在、すなわち、変異誘発の間に導入された第1突然変異により、最終変異体の発現の12.5%(8.4mg/L)に可溶性発現が増強された。A159Vバックグラウンドへの第2置換(L131P)の導入により、最終変異体の発現の42%(28mg/L)に可溶性発現が増強され、L131P、A159Vバックグラウンドへの第3置換(D38N)の導入により、67.2mg/Lに可溶性発現が増加した(図3)。野生型TrzNバックグラウンドにおいて、L131P突然変異は、溶解性においてA159V置換と類似の効果を有した(6.6mg/L収率)。したがって、突然変異を組み合わせることの効果は、相加的というよりむしろ相乗的である。実際に、D38N、L131変異体は、L131P変異体よりかなり低い可溶性であったが、D38N変異体の収率は、野生型TrzNについて以前に報告された収率と類似しており、このことは、A159V置換が存在する場合を除き、D38N置換が、酵素収率にネガティブに影響することを示唆する。
【0193】
基質の範囲
サイズ排除クロマトグラフィーによるTrzN cc3.2の精製により、これは75から150kDaの間の天然の分子量を有することが明らかになったが、このことは、それがホモ二量体またはホモ三量体であることを示唆する。これは、ホモ六量体であるAtzAと対照的であり、TrzNは単量体であるという以前の報告と対照的である(Shapirら、2006)。
【0194】
精製した酵素を、反復3のTrzN変異体の基質範囲を決定するために使用した。TrzN cc3.2は、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンおよびシアノジンを加水分解できたが(表7)、これらは、ハロゲンおよびメチルチオール脱離基、N−エチル、N−イソルプロピル、N−ジエチルならびにN−シアノジメチルメチルアルキル側鎖を有するs−トリアジンを代表する。メチルチオールおよびメトキシ脱離基の化学的性質が高度に類似するため、TrzNcc3.2は、以前に報告された、メトキシ−s−トリアジン(例えば、アトラトン)に対する活性を保持することが期待される(Shapirら、2005b)
【0195】
【表7】
【0196】
宿主およびベクターの範囲
TrzNcc3.2および野生型TrzNの両方についての全体のコード領域を、誘導性の高レベルな発現ベクターpETcc2から、低レベルの恒常的な発現ベクター中に移動した(pCS150、Scottら、2009)。結果のベクターを、大腸菌JM109、DH10βおよびBL21λDE3細胞の形質転換に使用した。結果の6つの菌株を、アトラジン清澄化プレートアッセイを用いて、それらの清澄化速度について試験した。各場合において、TrzNcc3.2発現菌株は、37℃で2日以内に清澄化したが、野生型TrzNを発現する菌株は、37℃で12日後も清澄化しなかった。これは、TrzN発現の改良が、プラスミドまたは菌株のどちらの依存性でもなかったことを実際に示す。
【実施例2】
【0197】
増強した活性を有するTrzN突然変異体の構造
方法および材料
構造解析
データ収集の統計は、他で報告されている(Jacksonら、2006)。単一波長異常回折(SAD)(Dauterら、2002)を用いる金属酵素の活性部位の金属からの相決定は、以前に報告されている(Liuら、2005)。以前の仕事により、非対称性ユニットは、TrzN二量体(Jacksonら、2006)を含むことが示唆されたため、CCP4一式のプログラム(Collaborative−Computational−Research−Project−4、1994)において実行されるように、SHELXD(Schneiderら、2002)を、1.28269Aで収集されたSDAデータを使用する異常差異パターソン合成(anomalous difference Patterson synthesis)において、4つのZn2+イオン部位の位置を決めるのに使用した。MLPHARE(Otwinowski、1991)を、続いて、4つの部位の占有率を精密にし、2.43の相力を用いて初期相を得るために使用した。SHELXEは、密度修正および相の改良のために使用した(Sheldrickら、2002);高い溶媒含量(79%)は、疑いの余地なく相の質に寄与した(Terwilliger、2001)。
【0198】
モデル構築および精密化
初期相を、ARP−wARP(Perrakisら、2001)を用いる自動モデル構築を行うために使用した。これは、38.0%のRfreeを有する初期モデルを作製した。次いで相互作用的なモデル構築のいくつかのラウンドを、COOT(EmsleyおよびCowtan、2004)を用いて実行し、続いてREFMACv5.0(Murshudovら、1997)において実行するように構造の理想化をした後、Rfreeが28.7%になった。抑制した精密化および水分子の追加により、Rfreeは23.5%に減少した。次いでBファクターを、各鎖の残基1〜195、196〜255および256〜271を含む3つの剛体を用いて、TLS精密化(Winnら、2001)の20および10ラウンドにセットし、続いて最大尤度の精密化の3ラウンドにより、Rfreeは20.0%次いで19.6%までさらに低下した。TLSANL(Howlinら、1993)およびANISOANL(Winn、2001)を、TLS精密化によって生成されたデータを解析するために使用した。TLSANLによって生成されたリブレーションテンソル(libration tensor)を、1.5のスケール係数を用いて、RIBBONS(Carson、1991)を用いて可視化した。構造の幾何学的検証を、RAMPAGE(Lovellら、2003)を用いて行い、これによって全残基が、好都合なまたは許容される領域にあったことが示された;PROCHECK(Laskowskiら、1993)では、全立体化学パラメータが、より良いか正常の限界内であることを示し、SFCHECK(Vaguineら、1999)では、0.45A2のLuzattiプロットにより座標において全体のエラーを与えた。
【0199】
結果
TrzNcc3.2の構造を、サルガッソー海の環境試料から、27%同一の構造2PAJを用いて、分子の置換によって解析した(Argawal,R.ら、出版されていない)。正確な解析は、探索モデルの広範な「枝刈り」の後にのみ見出され、さらに次いで二量体の1つの分子のみを見出すことができ、これはプログラムPHASER(McCoyら、2007)を用いる2番目に良いヒットであった。二量体における2つ目の分子を、続いてプログラムMOLREP(VaginおよびTeplyakov、2000)を用いて見出した。53.4%のモデルに関する初期Rfreeは、広範なモデル構築および精密化の後、21.1%に減少した。
【0200】
TrzNの効果的に同一な49.6kDaの2つのサブユニット(図4)を含み、極角において、(θ、φ)=(93、90°)で非結晶性の2回回転軸2倍軸と関連する、結晶性非対称ユニットにおける99.8kDaの1つのホモ二量体が存在する。既知の構造との低い配列類似性にもかかわらず、TrzNcc3.2は、(β/α)8バレルの構造のフォールドを採用し、機能的多様性をもつ大きな金属依存性アミドヒドロラーゼスーパーファミリーに属する。2PAJなどの、タンパク質データバンク(PDB)におけるもっとも類似した構造は、ほとんどが未知の機能である。しかし、機能的注釈がされているもののうち、もっとも近い相対物は、21%配列同一性を有し、2.2Aの主鎖r.m.s.dである、サーモトガ・マリティマ由来のTm0938、5−メチルチオアデノシン/S−アデノシルホモシステイン/アデノシンデアミナーゼ(Hermannら、2007)、19%配列同一性を有し、2.5Aの主鎖r.m.s.dであるヒトグアニンデアミナーゼ(2uz9;Moche,Mら、出版されていない)、および19%配列同位性を有し、2.9Aの主鎖r.m.s.dである、枯草菌由来のイミダゾロンプロピオナーゼである(Yuら、2006)。
【0201】
TrzNの活性部位空洞(図5)は、各サブユニットの触媒ドメインの中央に位置する。基質のアトラジンがドッキングした活性部位アーキテクチャーの図は、図4に示される。ストランド2からのグルタミン残基に加えて、ストランド1(H63、H65)および5(H238)上に位置する、アミドヒドロラーゼモチーフの3つの保存されたヒスチジン、および推定上の水/ヒドロキシド求核試薬は、金属リガンドを構築する。活性部位のZn2+金属イオンは、三方両錐形で配位し、ここでH63、H65およびH238はエクアトリアルなリガンドを含み、Q142および水分子は軸のリガンドを含む。Q129との結合長(3.5A)は、予想したより長いが、この残基の最小の移動によって顕著に短くすることができた。Zn2+金属イオンが、低い占有率(約20%)で結合するときのみ、我々は、完全に占領された活性部位が、より強いQ129−Zn2+相互作用を有し得ることを示唆する。金属の低い占有率が、電子密度に基づくその決定を曖昧とするため、ZnのK−エッジで収集した異常データを、金属酵素において活性部位の金属イオンを同定するのに効果的であることが示されている、Zn2+−TrzNの活性部位のBijvoet差異フーリエマップを計算するために使用した。このマップは、Zn2+が、予想した位置において活性部位で結合することを明白に示した。求核性の水は、アミドヒドロラーゼモチーフの保存された残基であるH274と付加的に水素結合し、ひいてはD300と水素結合して、H−D触媒性二分子を形成することが示される。異なるpH値での動力学的解析により、求核試薬は、溶液中で8.4のpKa値を有する、Zn2+−OH−のさらなる活性化と一致する、およそ8のpKaを有することが示唆される(図9)。
【0202】
一見、金属イオン依存性アミドヒドロラーゼファミリーの他のメンバーの活性部位と類似しているが、1つの大きな違いがある:他の全アミドヒドロラーゼ構造に存在する保存されたアスパラギン酸金属リガンドが、金属イオンリガンドとして働けないスレオニン残基と置き換わっている。アスパラギン酸は、類似しない位置でQ129と明らかに機能的に置き換わっており、これによって金属イオンの三方両錐配位形が回復される。荷電金属イオンリガンドの極性リガンドによる置換は、金属イオン親和性を低下させる可能性がある(図7)。エンテロバクター・アエロゲネス由来の金属ホスホジエステラーゼの活性部位は、この点において情報を与える;活性部位における2つの金属イオンの配位圏は、α部位におけるアスパラギン酸の、β位置におけるアスパラギンによる置換が異なり、これによってβ部位に対する金属イオン親和性の著しい低下が生じることが示された。実際に、TrzNも、金属イオンに対する相対的に弱い親和性を有するようである;過剰なZn2+を増殖培地に添加し、金属キレート剤を、精製または結晶化の間に使用しなかったが、結晶構造は、亜鉛の非常に低い占有率を有する(上記を参照されたい)。
【0203】
Zn2−についての解離定数を、多くのZn2+含有酵素について決定されるものより5から6オーダーの間のマグニチュードで高い、2.6μMであると決定し(図6)、これは、しばしば低いピコモーラー範囲で亜鉛についてKdを有する(炭酸脱水酵素Kd=4pM;グリオキシラーゼI、Kd=27pM;ジペプチジルペプチダーゼIII、Kd=17pM;カルボキシペプチダーゼA、Kd=1.6pMおよびスーパーオキシドジスムターゼ、Kd=10pM。
【0204】
TrzNの基質結合ポケットを特性評価するために、アトラジンを手動で活性部位中にモデリングした(図5および7)。空洞を、4つの区切りに分けることができる;イソプロピルおよびエチル側鎖ポケット、アトラジンの芳香環とπ−πスタッキングにより相互作用する残基、ならびに基質および/または産物と水素結合する残基。金属イオンリガンドH238に加えて、イソプロピル側鎖ポケットは、M82、L86、P131、F132、M163、C198およびY215の側鎖によって形成される。エチル側鎖ポケットは、求核試薬リガンドH274ならびに4つの残基の側鎖P299、D300、M303およびW305によって形成される。ポケットの「ベース」で、W85は、アトラジンの芳香環とπ−πスタッキング相互作用を形成し、これによって結合および遷移状態の間に発生する負電荷の両方を安定化することになる。最後に、E241は、カルボキシル基の酸素原子が、イソプロピル側鎖のNH基から3.3Aであるとき、アトラジンと水素結合を形成するように位置するが、2.9A水素結合によってリンクしたS328−T325二分子は、加水分解によって産生される塩素イオンと相互作用できるようになり、4面体型中間体上に発生する負電荷を安定化する。
【0205】
興味深いことに、ノカルディオイデス菌種株AN3由来の天然TrzN変異体における最近の研究により、アメトリンなどの乏しい脱離基を有する基質に対する触媒活性を消滅させるE241Q突然変異が同定された(Yamazakiら、2008)。本発明者等らは、我々の系におけるこれらの影響を試験し、この突然変異の結果として活性が変化することも発見したが、結合(Km)というよりむしろ主に基質のターンオーバー(kcat)の低下の結果であることを示唆する(表8)。ターンオーバー速度の影響を受けるという観察は、乏しい脱離基を有する基質との低下した活性を有するQ241突然変異体とも一致し、これは、突然変異により、反応への活性化エネルギーを効果的に低下させる酵素の能力が低下されることを示唆する。同じ報告において、位置214および215は、乏しい基質への触媒作用に影響を与えることも確認された。再び我々は、これがkcatの低下によることを示す(表8)。全体的に見て、基質結合性空洞の相補的な性質は、形状および疎水性の両方に関して際立っている。基質/産生成物の水素結合に関わる金属リガンドおよび残基を除き、11/12残基は疎水性であり、同等に疎水的なアトラジン基質の密接でエネルギー的に好都合な結合および脱溶媒和を促進することになるであろう。
【0206】
【表8】
【0207】
活性部位がアトラジンに理想的に適合するようであるが、それはバルク溶媒に完全に閉じ、すなわちアトラジンがこの基質結合ポケットに入ることができる道がないことは興味深い。したがってこれは、立体構造動力学によりが、いくらかの信頼を有もってする基質ターンオーバーに不可欠であることを示唆できる事例である。閉じた活性部位間隙は、図5および7に示し、疎水性残基(L86、M92、L172、Y215、L243、M247、M303およびW305)のネットワークは、活性部位の入口で相互作用し、閉じたそれを効果的に「ジッピング」して閉じる。「ロック」は、ループ3のL172であり、これはネットワークのトップに位置し、定位置に他の残基を効果的に保持することになるであろう。活性部位が開くもっともらしい機構は、特定の可動性ループの尖部に位置するL185に主に関わり、ここで残りのタンパク質に関連する平均的なBファクターが、部分的な占有率/高い可動性と非常に高度に一致する。このループにおける立体構造の変化は、活性部位間隙の2つの側を解開放して、同様の酵素で観察されている「ゆらぎ」の挙動で分離し(Jacksonら、2007)、基質が入り、産生成物が出ていくのを可能にするようである。実際に、このファミリーのメンバーにおける表面ループの立体構造の変動の役割は、最近報告されており、触媒作用(閉じる)および拡散(開く)について最適化した立体構造の基質の間で酵素を切り換えることによって、ターンオーバー速度をモジュールする働きをすることができた。
【0208】
TrzNの構造、基質のドッキングおよび動力学反応速度により、触媒機構を、図8に概説したように提案することができる。反応を4つのステップ(i)基質の結合および求核試薬性の産生、(ii)求核攻撃、(iii)4面体型中間体の分解、(iv)産生成物の放出に分けることができる。触媒作用において重要な役割を果たすようであるTrzNの活性部位に存在する2つの触媒性二分子がある。これらのうちの1つ目、D300−H274二分子は、活性部位の亜鉛金属イオンと協調して、求核性水酸化物の産生に必須のようである。活性部位のZn2+イオンは、結合水のpKaを低下させるルイス酸として働くことになる一方、D300により位置づけられ、安定化したH274は、その脱プロトン化に寄与して求核性水酸化物を形成するためのその脱プロトン化に寄与することになるであろう。基質の結合は、主にW85とのπ−πスタッキング相互作用およびE241との水素結合に関わる。乏しい脱離基に対する活性の損失(Yamazakiら、2008)およびこの残基のQ241への変異誘発におけるkcat数の減少(表8)は、E241との静電気的相互作用が、配向の最適化配向に加えて、基質を活性化する働きができることを示唆する。基質の結合および求核試薬性の産生の後、求核攻撃がC4炭素で起こることになり、非局在性の負電荷が新しいヒドロキシル基、塩素原子、および遷移状態の芳香トリアジン環内にわたって分散した非局在性の負電荷とともに、4面体型中間体の形成を引き起こす(アラインメントは不可能なSN2置換を不可能にする)。この電荷は、ヒドロキシル基でD300−H274二分子、塩素原子でS329−T325二分子、および芳香環でW85とのπ−πスタッキングによって安定化することになるであろう。この中間体の分解により、遊離クロライドに加えて、脱塩素化された産生成物ヒドロキシアトラジンが産生されることになるであろう。産生成物の放出には、次いで酵素における立体構造変化および活性部位間隙の開口が必要になる。
【0209】
結論として、以下の残基:M82、W85、L86、M92、P131、M163、L172、C211、Y215、H238、E241、L243、M247、H274、P299、D300、M303、W305、T325またはS329を、特異的活性、触媒定数(kcat)、基質特異性(Km)、安定性および/または二次速度定数(kcat/Km)を変えるために変化させることができる。
【実施例3】
【0210】
実地試験
方法および材料
ホモジネートを湯汲むTrzNcc3.2を含むホモジネートの調製
活性TrzNcc3.2を含む清澄化した細菌のホモジネートを、最少培地(10.6g/LのKH2PO4、4g/Lの(NH4)2HPO4、1.7g/Lのクエン酸一水和物、31.3mL/Lのグリセロール)で増殖した、TrzNcc3.2を発現するBL21λDE3の発酵物2リットルから調製した。10mL/LのPTM4塩(0.2g/LのD−ビオチン、2.0g/LのCuSO4.5H2O、0.08g/LのNaI、3.0g/LのMnSO4.H2O、0.2g/LのNa2MoO4、0.02g/Lのホウ酸、0.5g/LのCoCl2.6H2O、7.0g/LのZnC1l2、22.0g/LのFeSO4.7H2O、0.5g/LのCaSO4、1mL/LのH2SO4)をオートクレーブ高圧滅菌した後、0.6g/LのMgSO4を添加した。発酵に、アンピシリン150mg/Lおよびチアミン331mg/Lを補充したグリセロールを与え、IPTG11.9mg/Lで誘導した。発酵により、約240gの湿重量の細胞ペレットが産生された(OD600=122)。細胞をpH6.9のMOPS5.2g/L中で懸濁し、次いでホモジナイザーに3回通し、遠心分離によって清澄化した。清澄化したライセートを、0.22μMのフィルターに通して、インタクトな細胞を除去し、DNaseIをインタクトなDNAの除去に使用した。酵素活性を、Souzaら(1996)により記載されたUV光吸収法およびScottら(2009)に記載された比色法の両方を用いて決定した(アトラジン258±19mg/mgライセート/分)。ホモジネートを−80℃で保存し、必要な時に4℃で解凍した。
【0211】
試験ダムの調製
オーストラリアのクイーンズランドの湿潤/乾燥熱帯のクレア近くのサトウキビ農場(緯度19:48、経度147:14)で、約1.5MLの貯水ダムを、所要量のアトラジン(ヘクタールあたり3.3kg)で前処理した農地の灌漑からの源流で満たした。細菌のホモジネート240gを水20リットル中で懸濁し、貯水ダムの表面にわたって均一に広げることにより散布した。ダムをアトラジン汚染流出水で満たす前、酵素を添加する前、および酵素を添加した後の時間間隔で、ダムをアトラジン含有流出水で満たす前に、1リットルの試料2組を採取した。酵素反応を止めるために試料をすぐに氷上に保存した。氷上に置いて4時間以内に試料を凍結させた。
【0212】
アトラジン濃度の決定
アトラジン濃度を、直接注入投与を用いるように修正した、Lewisら(2009)に記載のLCMSMS法、および以下のLCMS法によるCSIRO Entomologyによって、2つの独立した実験室;Queensland Health Forensic and Scientific Services(QHFSS)で決定した。簡潔には、試料100mLを、HClを用いてpH2.8に酸性化し、次いで試料中のアトラジンを、事前に条件付けしたOasis SPE Max Cartridges(Waters、USA)を用いて、固相抽出によって1000倍濃縮し、(アンモニアを含む)MeOH3mL中に溶出した。試料を続いて乾燥させ、MeOH100μl中で溶解した。試料をHPCLによって分離し、265nmでの吸光度を測定することによってアトラジン濃度についてアッセイし、分析物のピーク領域を、Analystソフトウェアを用いて計算した。増幅させた試料は、10%以内の一致であった。HPLCピークの同一性を、質量スペクトル分析によって確認し、それによってアトラジンイオン216m/zを、Agilent ToF−MSD上に抽出した。
【0213】
結果
オーストラリアのクイーンズランドの湿潤/乾燥熱帯のクレア近くのサトウキビ農場(緯度19:48、経度147:14)で、約1.5MLの貯水ダムを、所要量のアトラジン(ヘクタールあたり3.3kg)で前処理した農地の灌漑からの源流で満たした。TrzNcc3.2を産生する細菌のホモジネート240gを、水20リットル中で懸濁し、貯水ダムの表面にわたって均一に広げることによって手で適用アプライした。ダムをアトラジン汚染流出水で満たす前、酵素を添加する前、および酵素を添加した後の時間間隔で、ダムをアトラジン含有流出水で満たす前に、1リットルの試料2組を採取した。酵素反応を止めるために試料をすぐに氷上に保存した。わずか4時間氷上に置いて4時間以内に後、試料を凍結させた。
【0214】
灌漑の放出された水を採取する前、貯水ダム中の水は、アトラジン8〜12μg/Lを含んだ(データは示さない)。灌漑の放出された水で満たした後、アトラジン濃度は、157〜170μg/Lに上昇した(図10)。酵素の添加後、アトラジンの枯渇速度にずれがあり、これは酵素を貯水ダム中の水と混合した速度にもっとも起因するようであった。このやり方で散布した酵素についての「混合相」の持続時間は、レメディエーションされる水体の、体積や、表面積:体積の割合にほぼ確実に依存し、すなわち、より大きな水体や、小さな表面積:体積の割合を有する水体はより長期の混合相を必要とするであろう。
【0215】
混合相の間のずれにもかかわらず、酵素の添加は、添加後、最初の4時間に、アトラジンの濃度における>90%枯渇を導いた。この結果は、トリアジンについてのTrzNベースのバイオレメディエーションが、技術的に実行可能であることを示す。
【0216】
広く記載した本発明の精神または範囲から逸脱することなく、特定の実施形態において示した本発明に、多数の変形および/または改変を行うことができることが、当業者によって理解されるであろう。したがって、本実施形態は、あらゆる点で例示的であり、限定的ではないと見なされるべきである。
【0217】
本願は、US61/094,044からの優先権を主張し、その内容全体は、参照により本明細書に組み込まれている。
【0218】
本明細書で論じ、かつ/または参照したすべての刊行物は、その全体が本明細書に組み込まれている。
【0219】
本明細書に含めた文献、行為、物質、デバイス、物品などの任意の考察は、単に本発明についての背景を提供する目的のためである。任意またはすべてのこれらの事柄は、従来技術の基礎の一部を形成する、またはこれが本願の各請求項の優先日前に存在した場合、本発明に関連した分野における共通の一般的な知見であったことを認めるものと解釈されるべきでない。
【0220】
(参考文献)
【表9−1】
【表9−2】
【0221】
配列表への鍵配列番号1−野生型TrzNのアミノ酸配列
配列番号2−TrzNをコードする、コドンを最適化したオープンリーディングフレーム(TrzNco)
配列番号3−TTTからTTCへの156番目のコドン変化を有するTrzNをコードする、コドンを最適化したオープンリーディングフレーム(Trz L1)
配列番号4−野生型TrzNをコードするオープンリーディングフレーム
配列番号5から30−オリゴヌクレオチドプライマー
【特許請求の範囲】
【請求項1】
s−トリアジンおよび/またはジアジンを加水分解するポリペプチドをコードする、単離された、かつ/または外因性のポリヌクレオチドであって、前記ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリヌクレオチド。
【請求項2】
配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項3】
以下のヌクレオチド置換の1つもしくは複数、またはそこに対応するヌクレオチド位置での置換を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む、請求項1または請求項2に記載のポリヌクレオチド;T5C、C39A、C76A、C84A、T87C、C101A、T108A、T108A、G112A、A127G、C135T、A157T、C165T、G168A、C180T、C189T、A200T、C207T、G210A、G225T、A228G、C229T、T240C、A250C、C268A、G270A、A271T、T273A、C279T、A296G、A302G、A303G、A314G、C315A、T317C、T320C、A326C、A333G、T336C、C346T、G357A、A367G、C372T、C375A、C381T、T384C、C391A、C391G、C391T、T392C、T392A、T392G、G393C、T399C、C410T、C411A、C411T、A414G、A418C、T423C、T426A、A432T、C438T、C449G、C454T、T466C、T468C、T471C、C474T、G475A、C476T、A481G、C483T、G489A、G489T、T498C、T531A、A537G、A540G、A545G、T546G、G548A、T555C、T555C、C564T、G567A、G567A、G568A、G569A、G573C、T579C、A584T、G589A、A600G、T618C、T618C、T627C、C628G、G630A、C633T、G637A、T639C、T639C、A654G、G660A、G660T、T663C、C675A、G681T、C686T、C690A、C696T、G705A、A723G、C727G、T728C、T728G、G729C、G736A、G737C、G737A、G737T、T738G、T738C、T738C、G745A、G753C、G768A、T774A、C807A、T840A、A843G、A852T、A855G、C867T、T879C、G880A、G880T、G880C、C881T、G882T、C885T、G897A、T900C、T906A、A928G、A938T、T941C、C957A、T959A、C972T、T978A、C981T、C993T、C999T、C1003A、C1003T、T1011C、G1048A、G1048T、G1048C、A1049T、A1049G、G1053A、A1059G、A1086G、G1094A、T1101C、T1101G、C1128T、A1152G、G1176T、C1186A、C1186T、T1196C、C1203T、G1221A、C1223T、C1236T、G1248T、G1270A、C1278T、T1286A、T1305C、G1309A、C1321T、A1326G、C1329T、C1329T、C1332T、C1344A、C1351AおよびG1353T。
【請求項4】
以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする、請求項1から3のいずれか一項に記載のポリヌクレオチド;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【請求項5】
配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含む、請求項1から4のいずれか一項に記載のポリヌクレオチド。
【請求項6】
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンもしくはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含むポリペプチドをコードする、請求項1から5のいずれか一項に記載のポリヌクレオチド。
【請求項7】
以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする、請求項1から5のいずれか一項に記載のポリヌクレオチド;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【請求項8】
以下のヌクレオチド置換もしくは置換グループの1つ、またはそこに対応するヌクレオチド位置(複数可)での置換(複数可)を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む、請求項1から5のいずれか一項に記載のポリヌクレオチド;
i)T468C、
ii)T468C、A938T、
iii)A200T、G210A、T468C、
iv)T468C、C476T、G753C、
v)T468C、C476T、T728C、
vi)T468C、1048T、
vii)T384C、T468C、G569A、G681T、
viii)T468C、G475A、
ix)C279T、T468C、C1223T、C1329T、
x)C76A、T468C、A481G、
xi)C39A、C101A、T468C、T639C、G737C、G1048T、
xii)C410T、A418C、T468C、A600G、
xiii)T468C、G705A、C1003A、
xiv)T468C、G573C、
xv)T468C、C474T、C628G、C1278T、
xvi)T399C、T468C、G880A、T900C、
xvii)T468C、C1236T、
xviii)T87C、A367G、T468C、
xix)T468C、G1176T、
xx)C135T、T468C、C1344A、
xxi)T468C、A852T、
xxii)T468C、T738C、
xxiii)C454T、T468C、
xxiv)A200T、T468C、G1309A、
xxv)T468C、G489T、G745A、
xxvi)C410T、T468C、
xxvii)T468C、G736A、
xxviii)G225T、C268A、A414G、T468C、T627C、C1321T、
xxix)A432T、T468C、T471C、C476T、T728C、G1053A、C1351A、
xxx)A303G、C449G、T468C、C476T、C686T、C690A、T728C、C1128T、
xxxi)A200T、G210A、C372T、T468C、A654G、C1003A、
xxxii)C180T、A200T、A302G、C375A、T399C、C411T、T468C、A540G、G880A、T900C、
xxxiii)C229T、T468C、G705A、C1003A、
xxxiv)T317C、T468C、A481G、T531A、G753C、T906A、T978A、C1003A、A1326G、
xxxv)A127G、T320C、T468C、C476T、G753C、G1048T、
xxxvi)A157T、C410T、A418C、T468C、A545G、G568A、A600G、C628G、G630A、G1048T、C1332T、
xxxvii)T468C、C476T、A723G、C1003A、G1048T、C1329T、
xxxviii)C84A、T399C、T468C、C483T、G880A、T900C、G1048A、
xxxix)T468C、T498C、C628G、C885T、A1086G、G1270A、
xl)T384C、T468C、C476T,G569A、G753C、A1152G、
xli)T336C、T468C、C476T、A537G、C564T、
xlii)A228G、T240C、T468C、C628G、G880A、G1048T、G1094A、
xliii)T273A、A367G、C381T、T399C、T468C、A481G、G880A、T900C、
xliv)A200T、C207T、G210A、T468C、C476T、G1048T、A1059G、
xlv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1236T、
xlvi)A200T、G210A、C346T、T468C、C476T、T579C、T728C、C1278T、
xlvii)C438T、T468C、C476T、A855G、
xlviii)G168A、T426A、T468C、C476T、C628G、C1203T、T1305C、
xlix)T468C、C476T、T663C、C1236T、
l)T384C、T468C、C476T、T728C、A928G、C1003A、C1186A、
li)T5C、C315A、T468C、C476T、G589A、G681T、T728C、G897A、C1003A、
lii)C279T、T468C、C476T、C1003A、
liii)A250C、A314G、T468C、C476T、
liv)A200T、G210A、T468C、T555C、G880A、T900C、
lv)T468C、C476T、T546G、C696T、C1003A、G1048T、
lvi)A200T、G210A、T468C、C476T、T728C、T1101G、
lvii)G112A、C165T、T468C、C476T、T618C、G753C、C999T、T1101C、
lviii)T423C、T468C、C476T、G489A、A584T、G753C、G1048T、
lix)T466C、T468C、C474T、C628G、G1048T、
lx)T108A、A200T、G210A、G357A、T468C、G1048T、A1059G、C1278T
lxi)T468C、C476T、G567A、G753C、G1048T、
lxii)A200T、G210A、A296G、T468C、C476T、G637A、T728C、C1003A、
lxiii)C189T、A326C、T468C、C476T、T941C、T959A、A1059G、C1186T、T1196C、G1248T、T1286A、
lxiv)T468C、C476T、C1003A、
lxv)A200T、G210A、C279T、T468C、C476T、T879C、C1003A、G1048T、
lxvi)G112A、C165T、T392C、T468C、C476T、T618C、G660A、C675A、G753C、C807A、C993T、C999T、T1101C、T1305C、
lxvii)A271T、T392C、T468C、C476T、G753C、G880A、G1048T、G1094A、C1186A、G1221A、
lxviii)A228G、T240C、T468C、G548A、C628G、G1048T、C1332T、
lxix)T108A、A200T、G210A、T423C、T468C、C476T、G567A、G1048T、
lxx)T384C、T468C、C476T、C628G、C867T、G880A、T900C、C981T、
G1048A、A1326G、
lxxi)A200T、C207T、G210A、T468C、C476T、T840A、T900C、G1048A、
lxxii)T468C、C476T、C633T、G660T、A723G、C1003A、G1048T、C1329T、
lxxiii)A228G、T240C、G270A、T468C、C628G、880A、G1048T、
lxxiv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1003T、C1236T、
lxxv)A228G、T240C、T468C、C628G、G880A、C957A、C1003A、または
lxxvi)A200T、G210A、C411A、T468C、T555C、T639C、A654G、G768A、T774A、C972T、C1003A、T1011C、G1353T。
【請求項9】
前記s−トリアジンが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンである、請求項1から8のいずれか一項に記載のポリヌクレオチド。
【請求項10】
前記細菌細胞が大腸菌である、請求項1から9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
配列番号2および/または配列番号4と少なくとも90%同一である配列を含む、請求項1から10のいずれか一項に記載のポリヌクレオチド。
【請求項12】
細胞中でポリヌクレオチドの発現を導くことができるプロモーターに作動可能に連結されている、請求項1から11のいずれか一項に記載のポリヌクレオチド。
【請求項13】
少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質をコードする、請求項1から12のいずれか一項に記載のポリヌクレオチド。
【請求項14】
請求項1から13のいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項15】
請求項1から13のいずれか一項に記載のポリヌクレオチドおよび/または請求項14に記載のベクターを含む宿主細胞。
【請求項16】
シャペロンをコードする外因性ポリヌクレオチドをさらに含む、請求項15に記載の宿主細胞。
【請求項17】
細菌細胞、酵母細胞または植物細胞である、請求項15または請求項16に記載の宿主細胞。
【請求項18】
請求項15から17のいずれか一項に記載の少なくとも1つの細胞を含むトランスジェニック植物。
【請求項19】
請求項15または請求項16に記載の少なくとも1つの細胞を含む、トランスジェニック非ヒト動物。
【請求項20】
s−トリアジンおよび/またはジアジンを加水分解する、実質的に精製された、かつ/または組換えポリペプチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリペプチド。
【請求項21】
配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含む、請求項20に記載のポリペプチド。
【請求項22】
以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む、請求項20または請求項21に記載のポリペプチド;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【請求項23】
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンもしくはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含む、請求項20から22のいずれか一項に記載のポリペプチド。
【請求項24】
以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含む、請求項20から22のいずれか一項に記載のポリペプチド;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【請求項25】
前記s−トリアジンが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンである、請求項20から24のいずれか一項に記載のポリペプチド。
【請求項26】
配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍のアトラジン加水分解活性を有する、請求項20から25のいずれか一項に記載のポリペプチド。
【請求項27】
少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質である、請求項20から26のいずれか一項に記載のポリペプチド。
【請求項28】
固体支持体に固定化されている、請求項20から27のいずれか一項に記載のポリペプチド。
【請求項29】
請求項15から17のいずれか一項に記載の宿主細胞、請求項18に記載の植物、および/または請求項19に記載の動物の抽出物であって、請求項20から28のいずれか一項に記載のポリペプチドを含む抽出物。
【請求項30】
請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、および/または請求項29に記載の抽出物、および1つまたは複数の許容される担体を含む組成物。
【請求項31】
s−トリアジンまたはジアジンを加水分解するための方法であって、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物と、s−トリアジンまたはジアジンを接触させるステップを含む方法。
【請求項32】
前記s−トリアジンまたはジアジンが、土壌、水、生体物質またはそれらの組合せからなる群から選択される試料中にある、請求項31に記載の方法。
【請求項33】
対象においてs−トリアジンまたはジアジンによって引き起こされた毒性を処理する方法であって、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物を対象に投与するステップを含む方法。
【請求項34】
対象においてs−トリアジンまたはジアジンによって引き起こされた毒性の処理用の薬剤を製造するための、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物の使用。
【請求項35】
s−トリアジンおよび/またはジアジンを加水分解できるポリペプチドを産生する方法であって、前記ポリペプチドをコードするポリヌクレオチドの発現を可能にする条件下で、前記ポリペプチドをコードする請求項15から17のいずれか一項に記載の宿主細胞、または前記ポリペプチドをコードする請求項14に記載のベクターを培養するステップと、発現したポリペプチドを回収するステップとを含む方法。
【請求項36】
宿主細胞を検出するための方法であって、
i)細胞または細胞集団を、細胞(複数可)によるポリヌクレオチドの取込みを可能にする条件下で、請求項1から13のいずれか一項に記載のポリヌクレオチドと接触させるステップと、
ii)ステップi)からの細胞またはその子孫細胞を、s−トリアジンまたはジアジンにさらすことにより宿主細胞を選択するステップと
を含む方法。
【請求項37】
前記ポリヌクレオチドが、請求項20から28のいずれか一項に記載のポリペプチドをコードする、請求項36に記載の方法。
【請求項38】
前記ポリヌクレオチドが、請求項1から13のいずれか一項に記載のポリヌクレオチドを含む第1オープンリーディングフレーム、および請求項1から13のいずれか一項に記載のポリヌクレオチドを含まない第2オープンリーディングフレームを含む、請求項36または請求項37に記載の方法。
【請求項39】
請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物を含む、s−トリアジンまたはジアジンを加水分解するためのキット。
【請求項40】
請求項20から27のいずれか一項に記載のポリペプチドの結晶。
【請求項41】
配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを設計する方法であって、請求項40に記載の結晶の原子座標を使用して、s−トリアジンまたはジアジンが候補ポリペプチドと結合する能力をコンピュータによって評価するステップと、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを選択するステップとを含む方法。
【請求項1】
s−トリアジンおよび/またはジアジンを加水分解するポリペプチドをコードする、単離された、かつ/または外因性のポリヌクレオチドであって、前記ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号2または配列番号4として提供されるヌクレオチド配列を含む外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリヌクレオチド。
【請求項2】
配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含むポリペプチドをコードする、請求項1に記載のポリヌクレオチド。
【請求項3】
以下のヌクレオチド置換の1つもしくは複数、またはそこに対応するヌクレオチド位置での置換を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む、請求項1または請求項2に記載のポリヌクレオチド;T5C、C39A、C76A、C84A、T87C、C101A、T108A、T108A、G112A、A127G、C135T、A157T、C165T、G168A、C180T、C189T、A200T、C207T、G210A、G225T、A228G、C229T、T240C、A250C、C268A、G270A、A271T、T273A、C279T、A296G、A302G、A303G、A314G、C315A、T317C、T320C、A326C、A333G、T336C、C346T、G357A、A367G、C372T、C375A、C381T、T384C、C391A、C391G、C391T、T392C、T392A、T392G、G393C、T399C、C410T、C411A、C411T、A414G、A418C、T423C、T426A、A432T、C438T、C449G、C454T、T466C、T468C、T471C、C474T、G475A、C476T、A481G、C483T、G489A、G489T、T498C、T531A、A537G、A540G、A545G、T546G、G548A、T555C、T555C、C564T、G567A、G567A、G568A、G569A、G573C、T579C、A584T、G589A、A600G、T618C、T618C、T627C、C628G、G630A、C633T、G637A、T639C、T639C、A654G、G660A、G660T、T663C、C675A、G681T、C686T、C690A、C696T、G705A、A723G、C727G、T728C、T728G、G729C、G736A、G737C、G737A、G737T、T738G、T738C、T738C、G745A、G753C、G768A、T774A、C807A、T840A、A843G、A852T、A855G、C867T、T879C、G880A、G880T、G880C、C881T、G882T、C885T、G897A、T900C、T906A、A928G、A938T、T941C、C957A、T959A、C972T、T978A、C981T、C993T、C999T、C1003A、C1003T、T1011C、G1048A、G1048T、G1048C、A1049T、A1049G、G1053A、A1059G、A1086G、G1094A、T1101C、T1101G、C1128T、A1152G、G1176T、C1186A、C1186T、T1196C、C1203T、G1221A、C1223T、C1236T、G1248T、G1270A、C1278T、T1286A、T1305C、G1309A、C1321T、A1326G、C1329T、C1329T、C1332T、C1344A、C1351AおよびG1353T。
【請求項4】
以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする、請求項1から3のいずれか一項に記載のポリヌクレオチド;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【請求項5】
配列番号2または配列番号4のヌクレオチド番号468に対応する位置にシトシンを含む、請求項1から4のいずれか一項に記載のポリヌクレオチド。
【請求項6】
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンもしくはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含むポリペプチドをコードする、請求項1から5のいずれか一項に記載のポリヌクレオチド。
【請求項7】
以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含むポリペプチドをコードする、請求項1から5のいずれか一項に記載のポリヌクレオチド;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【請求項8】
以下のヌクレオチド置換もしくは置換グループの1つ、またはそこに対応するヌクレオチド位置(複数可)での置換(複数可)を有する配列番号2または配列番号4として提供されるヌクレオチド配列を含む、請求項1から5のいずれか一項に記載のポリヌクレオチド;
i)T468C、
ii)T468C、A938T、
iii)A200T、G210A、T468C、
iv)T468C、C476T、G753C、
v)T468C、C476T、T728C、
vi)T468C、1048T、
vii)T384C、T468C、G569A、G681T、
viii)T468C、G475A、
ix)C279T、T468C、C1223T、C1329T、
x)C76A、T468C、A481G、
xi)C39A、C101A、T468C、T639C、G737C、G1048T、
xii)C410T、A418C、T468C、A600G、
xiii)T468C、G705A、C1003A、
xiv)T468C、G573C、
xv)T468C、C474T、C628G、C1278T、
xvi)T399C、T468C、G880A、T900C、
xvii)T468C、C1236T、
xviii)T87C、A367G、T468C、
xix)T468C、G1176T、
xx)C135T、T468C、C1344A、
xxi)T468C、A852T、
xxii)T468C、T738C、
xxiii)C454T、T468C、
xxiv)A200T、T468C、G1309A、
xxv)T468C、G489T、G745A、
xxvi)C410T、T468C、
xxvii)T468C、G736A、
xxviii)G225T、C268A、A414G、T468C、T627C、C1321T、
xxix)A432T、T468C、T471C、C476T、T728C、G1053A、C1351A、
xxx)A303G、C449G、T468C、C476T、C686T、C690A、T728C、C1128T、
xxxi)A200T、G210A、C372T、T468C、A654G、C1003A、
xxxii)C180T、A200T、A302G、C375A、T399C、C411T、T468C、A540G、G880A、T900C、
xxxiii)C229T、T468C、G705A、C1003A、
xxxiv)T317C、T468C、A481G、T531A、G753C、T906A、T978A、C1003A、A1326G、
xxxv)A127G、T320C、T468C、C476T、G753C、G1048T、
xxxvi)A157T、C410T、A418C、T468C、A545G、G568A、A600G、C628G、G630A、G1048T、C1332T、
xxxvii)T468C、C476T、A723G、C1003A、G1048T、C1329T、
xxxviii)C84A、T399C、T468C、C483T、G880A、T900C、G1048A、
xxxix)T468C、T498C、C628G、C885T、A1086G、G1270A、
xl)T384C、T468C、C476T,G569A、G753C、A1152G、
xli)T336C、T468C、C476T、A537G、C564T、
xlii)A228G、T240C、T468C、C628G、G880A、G1048T、G1094A、
xliii)T273A、A367G、C381T、T399C、T468C、A481G、G880A、T900C、
xliv)A200T、C207T、G210A、T468C、C476T、G1048T、A1059G、
xlv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1236T、
xlvi)A200T、G210A、C346T、T468C、C476T、T579C、T728C、C1278T、
xlvii)C438T、T468C、C476T、A855G、
xlviii)G168A、T426A、T468C、C476T、C628G、C1203T、T1305C、
xlix)T468C、C476T、T663C、C1236T、
l)T384C、T468C、C476T、T728C、A928G、C1003A、C1186A、
li)T5C、C315A、T468C、C476T、G589A、G681T、T728C、G897A、C1003A、
lii)C279T、T468C、C476T、C1003A、
liii)A250C、A314G、T468C、C476T、
liv)A200T、G210A、T468C、T555C、G880A、T900C、
lv)T468C、C476T、T546G、C696T、C1003A、G1048T、
lvi)A200T、G210A、T468C、C476T、T728C、T1101G、
lvii)G112A、C165T、T468C、C476T、T618C、G753C、C999T、T1101C、
lviii)T423C、T468C、C476T、G489A、A584T、G753C、G1048T、
lix)T466C、T468C、C474T、C628G、G1048T、
lx)T108A、A200T、G210A、G357A、T468C、G1048T、A1059G、C1278T
lxi)T468C、C476T、G567A、G753C、G1048T、
lxii)A200T、G210A、A296G、T468C、C476T、G637A、T728C、C1003A、
lxiii)C189T、A326C、T468C、C476T、T941C、T959A、A1059G、C1186T、T1196C、G1248T、T1286A、
lxiv)T468C、C476T、C1003A、
lxv)A200T、G210A、C279T、T468C、C476T、T879C、C1003A、G1048T、
lxvi)G112A、C165T、T392C、T468C、C476T、T618C、G660A、C675A、G753C、C807A、C993T、C999T、T1101C、T1305C、
lxvii)A271T、T392C、T468C、C476T、G753C、G880A、G1048T、G1094A、C1186A、G1221A、
lxviii)A228G、T240C、T468C、G548A、C628G、G1048T、C1332T、
lxix)T108A、A200T、G210A、T423C、T468C、C476T、G567A、G1048T、
lxx)T384C、T468C、C476T、C628G、C867T、G880A、T900C、C981T、
G1048A、A1326G、
lxxi)A200T、C207T、G210A、T468C、C476T、T840A、T900C、G1048A、
lxxii)T468C、C476T、C633T、G660T、A723G、C1003A、G1048T、C1329T、
lxxiii)A228G、T240C、G270A、T468C、C628G、880A、G1048T、
lxxiv)A271T、A333G、T399C、T468C、C476T、A843G、G880A、T900C、C1003T、C1236T、
lxxv)A228G、T240C、T468C、C628G、G880A、C957A、C1003A、または
lxxvi)A200T、G210A、C411A、T468C、T555C、T639C、A654G、G768A、T774A、C972T、C1003A、T1011C、G1353T。
【請求項9】
前記s−トリアジンが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンである、請求項1から8のいずれか一項に記載のポリヌクレオチド。
【請求項10】
前記細菌細胞が大腸菌である、請求項1から9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
配列番号2および/または配列番号4と少なくとも90%同一である配列を含む、請求項1から10のいずれか一項に記載のポリヌクレオチド。
【請求項12】
細胞中でポリヌクレオチドの発現を導くことができるプロモーターに作動可能に連結されている、請求項1から11のいずれか一項に記載のポリヌクレオチド。
【請求項13】
少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質をコードする、請求項1から12のいずれか一項に記載のポリヌクレオチド。
【請求項14】
請求項1から13のいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項15】
請求項1から13のいずれか一項に記載のポリヌクレオチドおよび/または請求項14に記載のベクターを含む宿主細胞。
【請求項16】
シャペロンをコードする外因性ポリヌクレオチドをさらに含む、請求項15に記載の宿主細胞。
【請求項17】
細菌細胞、酵母細胞または植物細胞である、請求項15または請求項16に記載の宿主細胞。
【請求項18】
請求項15から17のいずれか一項に記載の少なくとも1つの細胞を含むトランスジェニック植物。
【請求項19】
請求項15または請求項16に記載の少なくとも1つの細胞を含む、トランスジェニック非ヒト動物。
【請求項20】
s−トリアジンおよび/またはジアジンを加水分解する、実質的に精製された、かつ/または組換えポリペプチドであって、ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドと少なくとも40%同一であり、
i)細菌細胞中で発現させると、配列番号1として提供されるアミノ酸配列をコードする外因性ポリヌクレオチドを含む同一条件下で培養した同系細菌細胞を用いる場合よりも多くのポリペプチドが産生され、かつ/または
ii)ポリペプチドが、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有する、ポリペプチド。
【請求項21】
配列番号1のアミノ酸番号159に対応する位置にスレオニンまたはバリンを含む、請求項20に記載のポリペプチド。
【請求項22】
以下のアミノ酸置換の1つもしくは複数、またはそこに対応するアミノ酸位置での置換を有する配列番号1に提供されるアミノ酸配列を含む、請求項20または請求項21に記載のポリペプチド;I2T、F13L、L26M、D28E、A34D、D38N、S43G、M53L、Y67F、S84R、L90M、T91S、D99G、K101R、D105E、D105G、V106A、I107T、E109A、I123V、L131P、L131N、L131T、L131D、L131V、L131G、L131C、L131S、L131Q、L131H、L131Y、L131I、T137I、S140R、T150S、F156L、A159T、A159V、S161G、M163I、F177L、D182E、D182G、R183H、G190D、G190S、Y195F、E197K、P210A、V213I、M227I、M227I、A229V、D230E、L243P、L243G、G246A、G246S、G246D、G246E、G246K、G246V、D249N、A294T、A294S、A294L、I310V、Y313F、L314P、V320E、L335M、D350N、D350Y、D350F、D350R、D350H、R365H、L396M、V399A、A408V、V424I、V429D、V437IおよびL451M。
【請求項23】
i)配列番号1のアミノ酸番号67に対応する位置にフェニルアラニン、および/または
ii)配列番号1のアミノ酸番号91に対応する位置にセリン、および/または
iii)配列番号1のアミノ酸番号131に対応する位置にプロリン、アスパラギン、スレオニン、アスパラギン酸、バリン、グリシン、システイン、セリン、グルタミン、ヒスチジン、チロシンもしくはイソロイシン、および/または
iv)配列番号1のアミノ酸番号159に対応する位置にスレオニンもしくはバリン、および/または
v)配列番号1のアミノ酸番号161に対応する位置にグリシン、および/または
vi)配列番号1のアミノ酸番号210に対応する位置にアラニン、および/または
vii)配列番号1のアミノ酸番号243に対応する位置にプロリンもしくはグリシン、および/または
viii)配列番号1のアミノ酸番号246に対応する位置にアスパラギン酸、セリン、グルタミン酸、リジン、バリンもしくはアラニン、および/または
ix)配列番号1のアミノ酸番号294に対応する位置にスレオニン、セリンもしくはロイシン、および/または
x)配列番号1のアミノ酸番号335に対応する位置にメチオニン、および/または
xi)配列番号1のアミノ酸番号350に対応する位置にチロシン、アスパラギン、フェニルアラニン、アルギニンもしくはヒスチジン、および/または
xii)i)からxi)のいずれか1つの生物学的活性断片
を含む、請求項20から22のいずれか一項に記載のポリペプチド。
【請求項24】
以下のアミノ酸置換もしくは置換グループの1つ、またはそこに対応するアミノ酸位置(複数可)での置換(複数可)を有する配列番号1に提供されるアミノ酸配列を含む、請求項20から22のいずれか一項に記載のポリペプチド;
i)Y313F
ii)Y67F
iii)A159V
iv)A159V、L243P
v)D350Y
vi)G190D、M227I
vii)A159T
viii)A408V
ix)L26M、S161G
x)F13L、A34D、G246A、D350Y
xi)T137I、S140R
xii)L335M
xiii)P210A
xiv)A294T
xv)I123V
xvi)Y67F、V437I
xvii)M163I、D249N
xviii)T137I
xix)G246S
xx)L90M
xxi)A159V、L243P、L451M
xxii)T150S、A159V、A229V、D230E、L243P
xxiii)Y67F、L335M
xxiv)Y67F、K101R、A294T
xxv)L335M
xxvi)V106A、S161G、F177L、L335M
xxvii)S43G、I107T、A159V、D350Y
xxviii)M53L、T137I、S140R、D182G、G190S、D350Y
xxix)A159V、L335M、D350Y
xxx)D28E、A294T、D350N
xxxi)P210A、V424I
xxxii)A159V、G190D
xxxiii)P210A、A294T、R365H、D350Y
xxxiv)I123V、S161G、A294T
xxxv)Y67F、A159V、D350Y
xxxvi)T91S、A159V、A294T
xxxvii)Y67F、A159V、L243P
xxxviii)A159V、P210A
xxxix)A159V、I310V、L335M、L396M、L243P
xl)I2T、D105E、A159V、E197K、M227I、L243P、L335M
xli)A159V、L335M
xlii)S84R、D105G、A159V
xliii)Y67F、A294T
xliv)A159V、D182E、L335M、D350Y
xlv)Y67F、A159V、L243P
xlvi)D38N、A159V
xlvii)A159V、M163I、Y195F、D350Y
xlviii)F156L、P210A、D350Y
xlix)Y67F、D350Y
l)A159V、D350Y
li)Y67F、D99G、A159V、V213I、L243P、L335M
lii)E109A、A159V、L314P、V320E、V399A、V429D
liii)A159V、L335M
liv)Y67F、A159V、L335M、D350Y
lv)D38N、L131P、A159V
lvi)T91S、L131P、A159V、A294T、R365H、L396M、D350Y
lvii)R183H、P210A、D350Y
lviii)Y67F、A159V、D350Y
lix)A159V、P210A、A294T、D350N
lx)Y67F、A159V、D350N
lxi)A159V、L335M、D350Y
lxii)P210A、A294T、D350Y
lxiii)T91S、A159V、A294T
lxiv)P210A、A294T、L335M、または
lxv)Y67F、L335M。
【請求項25】
前記s−トリアジンが、アトラジン、アメトリン、プロパジン、プロメトリン、シマジン、シメトリン、イパジン、トリエタジンまたはシアノジンである、請求項20から24のいずれか一項に記載のポリペプチド。
【請求項26】
配列番号1に提供されるアミノ酸配列を含むポリペプチドの少なくとも2倍のアトラジン加水分解活性を有する、請求項20から25のいずれか一項に記載のポリペプチド。
【請求項27】
少なくとも1つの他のポリペプチド配列をさらに含む融合タンパク質である、請求項20から26のいずれか一項に記載のポリペプチド。
【請求項28】
固体支持体に固定化されている、請求項20から27のいずれか一項に記載のポリペプチド。
【請求項29】
請求項15から17のいずれか一項に記載の宿主細胞、請求項18に記載の植物、および/または請求項19に記載の動物の抽出物であって、請求項20から28のいずれか一項に記載のポリペプチドを含む抽出物。
【請求項30】
請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、および/または請求項29に記載の抽出物、および1つまたは複数の許容される担体を含む組成物。
【請求項31】
s−トリアジンまたはジアジンを加水分解するための方法であって、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物と、s−トリアジンまたはジアジンを接触させるステップを含む方法。
【請求項32】
前記s−トリアジンまたはジアジンが、土壌、水、生体物質またはそれらの組合せからなる群から選択される試料中にある、請求項31に記載の方法。
【請求項33】
対象においてs−トリアジンまたはジアジンによって引き起こされた毒性を処理する方法であって、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物を対象に投与するステップを含む方法。
【請求項34】
対象においてs−トリアジンまたはジアジンによって引き起こされた毒性の処理用の薬剤を製造するための、請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物の使用。
【請求項35】
s−トリアジンおよび/またはジアジンを加水分解できるポリペプチドを産生する方法であって、前記ポリペプチドをコードするポリヌクレオチドの発現を可能にする条件下で、前記ポリペプチドをコードする請求項15から17のいずれか一項に記載の宿主細胞、または前記ポリペプチドをコードする請求項14に記載のベクターを培養するステップと、発現したポリペプチドを回収するステップとを含む方法。
【請求項36】
宿主細胞を検出するための方法であって、
i)細胞または細胞集団を、細胞(複数可)によるポリヌクレオチドの取込みを可能にする条件下で、請求項1から13のいずれか一項に記載のポリヌクレオチドと接触させるステップと、
ii)ステップi)からの細胞またはその子孫細胞を、s−トリアジンまたはジアジンにさらすことにより宿主細胞を選択するステップと
を含む方法。
【請求項37】
前記ポリヌクレオチドが、請求項20から28のいずれか一項に記載のポリペプチドをコードする、請求項36に記載の方法。
【請求項38】
前記ポリヌクレオチドが、請求項1から13のいずれか一項に記載のポリヌクレオチドを含む第1オープンリーディングフレーム、および請求項1から13のいずれか一項に記載のポリヌクレオチドを含まない第2オープンリーディングフレームを含む、請求項36または請求項37に記載の方法。
【請求項39】
請求項1から13のいずれか一項に記載のポリヌクレオチド、請求項14に記載のベクター、請求項15から17のいずれか一項に記載の宿主細胞、請求項20から28のいずれか一項に記載のポリペプチド、請求項29に記載の抽出物、および/または請求項30に記載の組成物を含む、s−トリアジンまたはジアジンを加水分解するためのキット。
【請求項40】
請求項20から27のいずれか一項に記載のポリペプチドの結晶。
【請求項41】
配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを設計する方法であって、請求項40に記載の結晶の原子座標を使用して、s−トリアジンまたはジアジンが候補ポリペプチドと結合する能力をコンピュータによって評価するステップと、配列番号1に提供されるアミノ酸配列を含むポリペプチドよりも高いs−トリアジンおよび/またはジアジン加水分解活性を有するポリペプチドを選択するステップとを含む方法。
【図1】

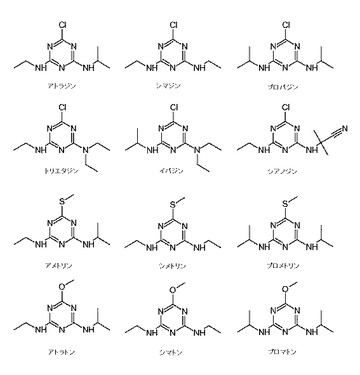
【図2】
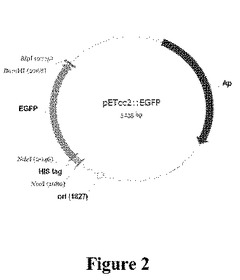
【図3】
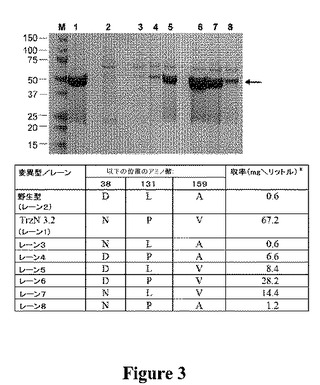
【図4】
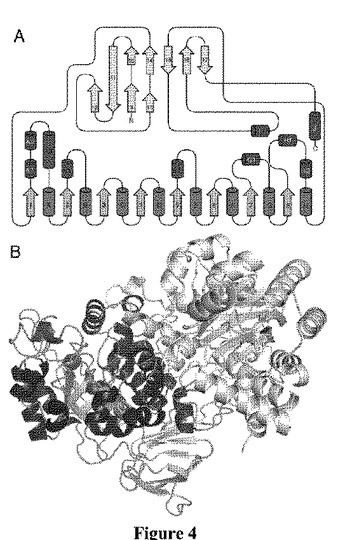
【図5】
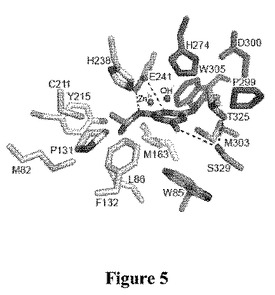
【図6】
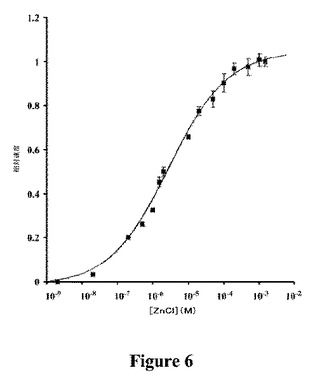
【図7】
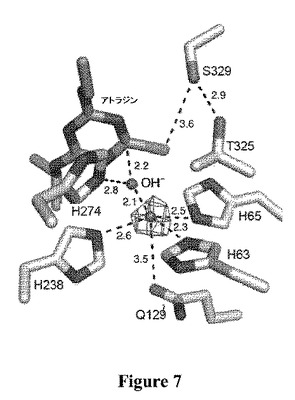
【図8】
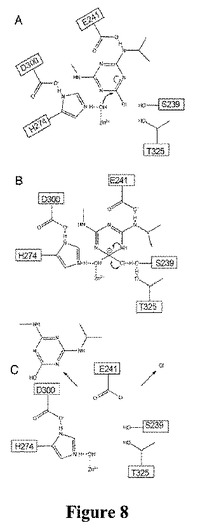
【図9】
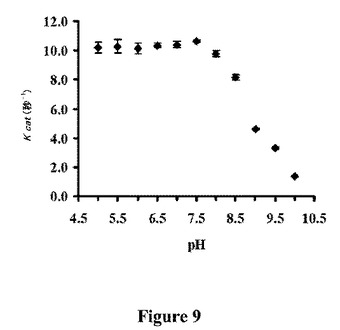
【図10】
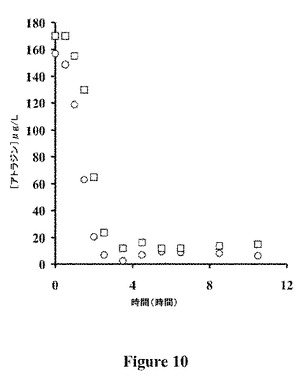
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2012−501628(P2012−501628A)
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願番号】特願2011−525373(P2011−525373)
【出願日】平成21年9月2日(2009.9.2)
【国際出願番号】PCT/AU2009/001136
【国際公開番号】WO2010/025499
【国際公開日】平成22年3月11日(2010.3.11)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
【公表日】平成24年1月26日(2012.1.26)
【国際特許分類】
【出願日】平成21年9月2日(2009.9.2)
【国際出願番号】PCT/AU2009/001136
【国際公開番号】WO2010/025499
【国際公開日】平成22年3月11日(2010.3.11)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【Fターム(参考)】
[ Back to top ]