
T細胞タンパク質チロシンホスファターゼの活性化方法およびそれに基づく治療方法
本発明は、T細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法および個体におけるチロシンキナーゼシグナル伝達の阻害方法に関する。本発明は、個体におけるチロシンキナーゼシグナル伝達を阻害することにより治癒可能である疾患または障害の予防または治療方法、およびT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化に基づく、個体における癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法に関する。本発明はまた、前記方法に有用な医薬組成物に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、T細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法および個体におけるチロシンキナーゼシグナル伝達の阻害方法に関する。さらに、本発明は個体におけるチロシンキナーゼシグナル伝達を阻害することにより治癒可能である疾患または障害の予防または治療方法に関する。さらにまた、本発明は、T細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化に基づく、個体における癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法に関する。本発明はまた、前記方法に有用な医薬組成物に関する。
【背景技術】
【0002】
本発明の背景を説明するために本明細書で使用する刊行物および他の資料、および特に、実施に関するさらなる詳細を提供するための事例は、引用により本明細書に組み込まれる。
【0003】
インテグリン媒介細胞接着は、増殖、生存および異なる細胞性シグナル伝達経路間のクロストークなどの多様な細胞応答を制御する1。これまでのところ、インテグリンは、足場依存性受容体チロシンキナーゼシグナル伝達を可能にする許可シグナルを伝えることが主に示されている2、3。細胞接着への応答において、インテグリンはある細胞質シグナル伝達経路を直接活性化し、間接的に成長因子受容体によるシグナル伝達を調節する。インテグリンの細胞質ドメインは、これらの機能を媒介するのに不可欠である4〜7。しかしながら、それらは固有の触媒活性を欠き、シグナル伝達のために細胞質タンパク質との相互作用を必要とする。コラーゲンは人体に非常に豊富に存在するタンパク質であるが、4つのコラーゲン結合インテグリンα1β1、α2β1、α10β1およびα11β1のシグナル伝達については相対的にほとんど知られていない。インテグリンα1β1は、コラーゲンとラミニンの受容体である。その1つのサブユニットは、カベオリン−1と関連していることが示されており、したがって、シグナル伝達アダプタータンパク質Shcを補充する(recruit)8。これは次にマイトジェン活性化タンパク質キナーゼ経路の活性化ならびにコラーゲン上の線維芽細胞の生存および増殖の増強をもたらす9。さらに、すべてのインテグリン細胞質テイルにみられる保存領域は、パキシリン、テーリン(talin)および焦点接着キナーゼとのつながりによって、焦点接着集合における機能が示されている10。しかしながら、これまでのところα1インテグリンにより明確に活性化されるシグナル伝達経路は確認されていない。
【発明の開示】
【0004】
本発明は、α−1−インテグリンの細胞質テイルが選択的かつ直接的に遍在的に発現するタンパク質チロシンホスファターゼTCPTP(T細胞タンパク質チロシンホスファターゼ)と相互作用し、コラーゲンへの細胞接着の際にそれを活性化させるという発見に基づく。その活性化により、EGF刺激の際のEGFR(上皮成長因子受容体)リン酸化が減少することとなる。α−1−インテグリンの細胞質テイルは、腫瘍抑制タンパク質、TCPTPの活性化を通してEGFRシグナル伝達の負の調節因子として機能する。α−1−細胞質ドメインペプチドの細胞への導入により、ホスファターゼ活性が誘導され、インビボでのヒト線維肉腫の接ぎ木(zenografts)においてはもとより、EGF誘導細胞増殖およびインビトロにおける悪性腫瘍の足場非依存性成長を誘導する。これらのデータは、インビボにおけるTCPTP活性の制御の最初の証明であり、受容体チロシンキナーゼシグナル伝達のインテグリン媒介性負調節の新規な分子パラダイムを表している。
【0005】
したがって、1つの側面によれば、本発明は、治療に使用するためのi)アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチド、またはii)哺乳類細胞において該ペプチドを発現可能なベクターのいずれかである薬剤に関する。
【0006】
もう1つの側面によれば、本発明は個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法であって、該個体に、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである有効量の薬剤を投与することによる方法に関する。
【0007】
第3の側面によれば、本発明は個体におけるチロシンキナーゼシグナル伝達の阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0008】
第4の側面によれば、本発明は個体における疾患または障害の予防または治療方法であって、該疾患または障害がチロシンキナーゼシグナル伝達を阻害することにより治癒可能なものであり、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0009】
第5の側面によれば、本発明は個体における、癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0010】
第6の側面によれば、本発明は、治療に効果的な量のTCPTPを活性化できる低分子またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれか、および薬学的に許容され得る担体を含む医薬組成物に関する。
【0011】
第7の側面によれば、本発明は、アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドをコードする核酸を包含する発現ベクターであり、哺乳動物細胞において該ペプチドを発現することができるベクター、および薬学的に許容され得る担体を含む医薬組成物に関する。
【発明を実施するための最良の形態】
【0012】
定義:
用語「治療」または「治療する」は、疾患または障害の完全な治癒ならびに該疾患または障害の改善または軽減を含むことを理解されたい。
【0013】
用語「予防」は、完全な予防(prevention)、防御(prophylaxis)、および前記疾患または状態を伴う病気になる個体のリスクを低下することを含むことを理解されたい。
【0014】
用語「個体」は、ヒトまたは動物の被験体を意味する。
【0015】
用語「有効量」は、特に動物またはヒトの被験体に投与する際に、所望の治療結果をもたらすのに充分な本発明の薬剤の任意の量を意味する。
【0016】
好ましい実施態様:
TCPTPの活性化に使用される薬剤は、TCPTPを活性化できる低分子であっても、またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれであってもよい。この配列は、α−1−インテグリンの細胞質テイルに特異的である。1つの実施態様によれば、ペプチドは正にアミノ酸配列RPLKKKMEK(配列番号1)であってよく、たとえば微量注入によって細胞に投与され得る。
【0017】
あるいは、ペプチドは、アミノ酸配列RPLKKKMEK(配列番号1)を包含するより長い鎖であってもよい。特に好ましい実施態様によれば、ペプチド(鎖RPLKKKMEK(配列番号1)またはそれを包含するより長い鎖)は、細胞膜透過性融合タンパク質の一部であってもよい。そのような融合タンパク質の例としては、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質転換ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む融合タンパク質があげられる。そのような融合タンパク質の例としては、YGRKKRRQRRRWKLGFFKRPLKKKMEK(配列番号2)があげられる。配列YGRKKRRQRRR(配列番号3)は、TATから誘導され、配列WKLGFFK(配列番号4)は、インテグリン(いずれのαインテグリンにも典型的な)から誘導される。
【0018】
さらなる代替によれば、投与される薬剤は、哺乳動物細胞において所望のペプチドを発現することができる核酸を含むベクターであってもよい。核酸はDNA配列中、RNA配列中またはウイルスベクター中に挿入することができる。そのようなウイルスベクターは、通常、アデノウイルス、アルファウイルス、アデノ随伴ウイルス、レトロウイルスまたはヘルペスウイルスに基づく。細胞におけるペプチドの発現については、たとえばM Parsonsら、Molecular and Cellular Biology, Aug. 2002, p.5897-5911.参照。
【0019】
よって、本発明によるTCPTPの活性化は、直接的または間接的に該活性化によって影響される障害または疾患を予防または治療するために使用され得る。
【0020】
少なくとも部分的に間接的に影響される疾患または障害の例としては、その治癒性がチロシンキナーゼシグナル伝達を阻害することにより恩恵を受け得る任意の疾患または障害があげられる。具体的には、そのような標的の非限定的な例は、上皮細胞成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼがあげられる。EGFRの阻害は、癌の予防、癌の成長、浸潤もしくは転移の予防または阻害に有用であり得る。しかしながら、これらの効果も、TCPTPの活性化による他の機序により部分的となり得るかもしれない。
【0021】
他方、EGFRの阻害はまた、他の、非癌疾患または障害の予防または障害に有用であり得る。
【0022】
インスリン受容体およびヤーヌスキナーゼの阻害は、インスリン受容体によって制御されるエネルギー代謝の異常を誘導するインスリン初期シグナル伝達の亢進と関係する疾患、ならびにインターフェロン受容体と結合するヤーヌスキナーゼの異常な活性を導くインターフェロンによる望ましくない炎症性シグナル伝達と関係する障害の治療または予防に有用であり得る。
【0023】
配列RPLKKKMEK(配列1)を含むペプチドは、細胞膜を通して透過できる形態でもたらされるのが好ましい。そのようなペプチドは、組成物の非経口投与に適した任意の担体と投与することができる。たとえば、ペプチドは脂質と複合化されるか、リポソームに詰められるか、シクロデキストリンもしくは他の複合化剤、生吸収性(bioresorbable)ポリマーまたは制御された放出投与のための他の好適な担体、ナノ粒子またはヒドロゲルに組み込まれることができる。
【0024】
あるいは、アミノ酸配列RPLKKKMEK(配列1)を含むペプチドをコードする核酸を包含する発現ベクターを個体に投与することもできる。核酸は、正に配列RPLKKKMEK(配列1)、またはこの配列を含むより長い配列をコードすることができる。ベクターは、たとえば脂質と複合化されるか、リポソームに詰められるか、シクロデキストリンもしくは他の複合化剤、生吸収性ポリマーまたは制御された放出投与のための他の好適な担体、ナノ粒子またはヒドロゲルに組み込まれることができる。
【0025】
そのような治療を必要とする患者に投与されるペプチドまたは発現ベクターの治療的有効量は、たとえば患者の年齢および体重、治療を必要とする正確な病状、およびその重篤度、および投与経路などの多数の因子に依存し得る。正確な量は、最終的には主治医の裁量であろう。したがって、本発明の実施は任意の用量、他の治療有効薬との組み合わせ、非経口投与のための薬学製剤またはデリバリーシステムを含む。ペプチドまたは発現ベクターは、全身的または局所的に投与される。適切な投与経路としては、静脈内、筋肉内、皮下注射、吸入、局所、眼、舌下、鼻、直腸、腹腔内デリバリー、およびイオン導入または他の経皮デリバリーシステムがあげられる。
【0026】
本発明を、以下の非制限的な実施例の項により説明する。
【実施例】
【0027】
方法
酵母ツーハイブリッドシステム
α1−インテグリン細胞質ドメインを、pGBKT7ベクター(クローンテック)におけるGAL4 DNA−BDに融合し、マウス17日胚適合マーカーcDNAライブラリー(BDクローンテック)をスクリーニングするためのベイトとして使用した。そのライブラリーのスクリーニングは、製造元のプロトコールにしたがって行なった。単離した独立したクローンの1つがGAL4−DBD−α1cytと相互作用したが、T細胞タンパク質チロシンホスファターゼをコードする無関係なベイトとは相互作用しなかった。
【0028】
タンパク質−タンパク質相互作用アッセイ
α1cytおよびα2cytをpGEX4T1(アマシャムバイオサイエンス)にサブクローニングした。全長TCPTP(45kD)およびその欠失変異体をpCGTC45プラスミド11からPCR増幅し、pGEX−6P−1にクローニングした。GST融合タンパク質をE.coli(BL21 pLysS)で発現させ、製造元(アマシャムバイオサイエンス)の取扱説明書にしたがい精製した。融合タンパク質をグルタチオンセファロースに固定し、25mMグルタチオンで溶出するか、またはGSTを製造元の取扱説明書にしたがって正確な(precession)プロテアーゼを用いて切断した。プルダウンアッセイについては、Hela細胞を溶解緩衝液(1%オクチルグルコシド、50mMトリス−HCl pH7.5、150mM NaCl、1mM MgCl2、完全プロテアーゼ阻害剤(ロッシュ))に溶解させ、遠心分離、10分、13000rpm+4Cで浄化(clarified)し、グルタチオンセファロースで前除去し(precleared)、そして固定化GST融合タンパク質とインキュベートし、同じ緩衝液1mlで3回洗浄した。結合したタンパク質をSDS−PAGEで分離し、ウエスタンブロットで検出した。あるいは、精製全長TCPTPを溶解緩衝液中で、合成インテグリンα1細胞質テイルペプチド(RPLKKKMEKPLKKKMEK(配列番号5)ジェネムド シンセシス)の存在下または非存在下で示すようにGST−融合タンパク質とインキュベートした。
【0029】
免疫沈降、ウエスタンブロッディングおよびホスファターゼアッセイ
血清枯渇(starved)Hela細胞をプラスチック上に未処理のまま置き、10%FBSで30分刺激するか、コラーゲンI型またはポリ−L−リジン(10μg/ml、シグマ)で被覆した組織培養プレート上に1時間置いた。示した場合、細胞は記述したように表面をビオチン化した12。細胞をLaemmli試料緩衝液に溶解させウエスタンブロット用にSDS−PAGE上で分画するか、または溶解緩衝液に溶解させ、各試料由来のタンパク質をそれぞれ等量Gタンパク質−セファロースで前除去し免疫沈降に付した。抗体(抗−TCPTP、オンコジーン;抗−a1、抗−a5、抗−a6、すべてケミコン;抗−SHP−2、サンタクルーズ)をGタンパク質−セファロースと共にインキュベートし、免疫沈降を4℃で2時間行なった。免疫沈殿物を細胞溶解緩衝液で3回洗浄した。共沈殿アッセイについては、タンパク質をSDS−PAGEで分離し、ウエスタンブロットで検出するか、Vectastain試薬(ベクター)でビオチンの検出を行なった。ホスファターゼアッセイについては、免疫沈殿物をホスファターゼ反応緩衝液(25mM Hepes pH7.4、50mM NaCl、1mM DTT)に再懸濁させ、反応物の1/3をウエスタンブロッディングに付し、残りのビーズを、基質としてdiFMUP(モレキュラープローブス)を用い、セリン/スレオニンホスファターゼ阻害剤カクテル(シグマ)の存在下で製造元の取扱説明書にしたがって三重でホスファターゼ活性を評価した。示した場合、合成ペプチドは、ホスファターゼアッセイ反応の前に15分間免疫沈殿物とプレインキュベートされた(α2ペプチドKLGFFKRKYEKMTKNPDEIDETTELSS(配列番号6)、Heino博士より提供)。精製タンパク質とのホスファターゼアッセイについては、全長TCPTPをホスファターゼ反応緩衝液で、示したようにTCPTP欠失変異タンパク質および合成インテグリン細胞質テイルペプチドの存在下または非存在下でインキュベートした。ウエスタンブロッディングをEGFR、EGFR(PY)845、992もしくは1068(セルシグナリングテクノロジーズ)、TCPTP(オンコジーン)またはチューブリン(サンタクルーズバイオテクノロジー)に対して作製された市販の抗体で行なった。抗体の特異的結合を過酸化水素共役二次抗体で検出し、拡張化学発光検出法により可視化した。
【0030】
トランスフェクション
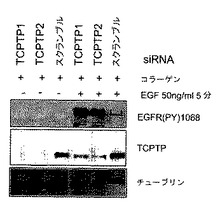
HT1080細胞を、以前に報告されているように13、Fugene6(ロッシュ)試薬を用いてpcDNA3.1/zeoベクター(バンダービルト大学、Pozzi博士より提供)中のα1cDNAで一時的にトランスフェクトした。TCPTPを標的とする2つの異なるアニールしたsiRNAs(ggcacaaaggaguuacauctt(配列番号7)、ggaguuacaucuuaacacatt(配列番号8);アンビオン(Anbion))またはスクランブル対照siRNAを、Hela細胞にオリゴフェクトアミン(インビトロジェン)を用い、製造元のプロトコールにしたがってトランスフェクトした。24時間後、トランスフェクション細胞を24時間血清枯渇させ、溶解およびウエスタンブロットアッセイの前に示したように処理した。
【0031】
免疫蛍光
I型コラーゲン、フィブロネクチンまたはポリ−L−リジン(10μg/ml)でコートした酸洗浄ガラスカバースリップ上に細胞を置き、1時間接着させた。細胞をPBSで洗浄し、4%パラホルムアルデヒドで10分間固定した。抗体染色については、細胞をPBS/0.1% Triton X−100に浸透させ(permeabilized)、ブロッキングのためにPBS/1%(w/v)BSAで洗浄した。細胞を室温で1時間、示した抗体で染色した。ついで、Alexa−488またはAlexa−555で共役された抗ウサギまたは抗マウス抗体(モレキュラープローブス)による検出の前に、細胞をPBSで3回洗浄した。PBSと水とで洗浄後、細胞をMowiol(100mM Tris−HCl pH8.5、10%(w/v)Mowiol(カルビオケム、サンディエゴ、CA)および25%(v/v)グリセロール含有アンチフェイド(antifade)(2.5%(w/v)1,4−ジアザジシクロ−2.2.2−オクタン(シグマ)))に載せた。スライドは、ツァィス倒立蛍光顕微鏡(Zeiss inverted fluorescence microscope)または共焦点レーザー顕微鏡(Axioplan 2 LSM 510;カールツァィス社(Carl Zeiss Inc.)、イェーナ、ドイツ)(63×/1.4 Plan−Apochromat油浸対物レンズを装備)を用いて調べた。共焦点画像は、約1.0μmの単一z−断面を表す。PBS中、1mM FDP(モレキュラープローブス)および1mg/mlのペプチドによる細胞質微量注入法を、ガラスの底3cmプレート上で培養されるHela細胞に行なった。生細胞の蛍光画像は、同一の設定で撮り、強度はZeiss Axioplan 4ソフトウエアを用いて定量化した。
【0032】
細胞増殖および軟寒天アッセイ
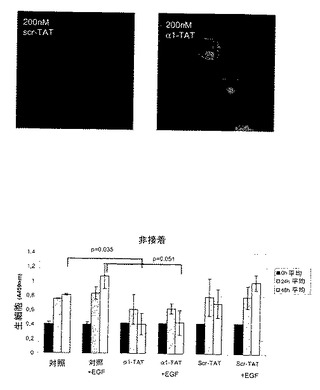
96ウェルプレートをBSAまたは示したマトリックスでコートした。細胞をそのウェルに104/ウェルで播いた。TATペプチド(α1−TAT fitc−YGRKKRRQRRRWKLGFFKRPLKKKMEK(配列番号2)、Scr−TAT fitc−YGRKKRRQRRRLKGWRFKLKPKFKEMK(配列番号2);ジェネムドシンセシス)およびEGF(シグマ)を示したように添加し、生存細胞の数をWST−1(ロッシュ)で製造元のプロトコールにしたがって検出した。軟寒天アッセイは記載されたように行なった14。200nM TATペプチドを含む培地を48時間毎に置き換え、9日間のインキュベーション後、培養物を二重盲検で撮影した。各視野(4×倍率)由来のコロニーの数と大きさをシンジーン(Syngene)のGeneToolsソフトウエアを用いて分析した。コロニーを大きさで分類した(小=200〜499ピクセル、中=500〜1000および大1000以上)。
【0033】
マウスおよび腫瘍細胞播種
雄の無菌ヌードマウスを4週齢で使用した。HT1080腫瘍細胞(5×106細胞/100μl PBS)をヌードマウスの背面右脇腹に皮下注射した。マウスを腫瘍細胞播種部位に隣接した皮下注射(20μl PBS、PBS中の20μM a1−TATまたは20μM Scr−TATペプチド)で、実験期間中週に3回処置した。播種後4週間でマウスを犠牲死させた。腫瘍を取り出し、その大きさを定量し、免疫組織化学用にパラフィンブロック中に加工した。IHCを以前に記載されたように行なった(Grant D.S.ら、Int. J. Cancer 2003 104: 121-129)。
【0034】
結果:
コラーゲンへのα1インテグリン−媒介細胞接着によって活性化されるシグナル伝達経路を特徴付けるために、本発明者らは、α1細胞質ドメイン(α1cyt)と相互作用するタンパク質を同定するために酵母ツーハイブリッドシステムを使用した。陽性クローンの1つは、TCPTPの45kDa型をコードしていた15。この分子は、遍在的に発現される核非受容体タンパク質チロシンホスファターゼであり、マイトジェンの刺激に応答して細胞質に移行することができる11。
【0035】
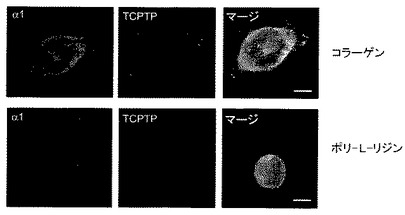
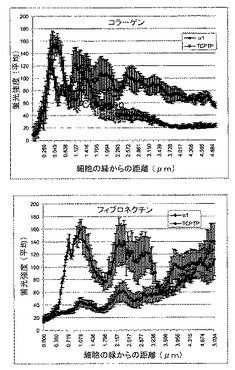
ヒト細胞における新規な相互作用を実証するために、本発明者らは、まず内因性α1インテグリンとTCPTPとが同じ場所に局在化する(colocalize)かどうかを調べた。PC3およびHeLa細胞を種々のマトリックスタンパク質上に置き、a1−インテグリンおよびTCPTPを二色免疫蛍光染色により可視化した。インテグリンを活性化できないポリ−L−リジン上に置かれた細胞では、TCPTPは細胞質および核の両方で見られた。4つのコラーゲン結合インテグリン以外のインテグリンに結合するフィブロネクチン上に置かれた細胞では、TCPTPは細胞質ゾルに広く見られた(図1B)。対照的に、コラーゲンへの結合では、TCPTPは、細胞膜の末梢領域においてα1β1インテグリンと同じ場所に局在化した(図1A、B)。これらのデータは、内因性インテグリンα1とTCPTPとがコラーゲンへの細胞接着により特異的にヒト細胞において同局在化することを示す。
【0036】
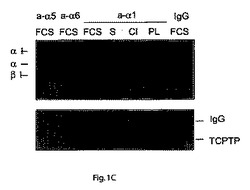
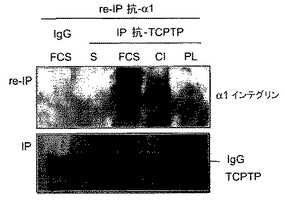
以前の研究では、TCPTPが分子的には特徴付けされていない機序によりマイトジェン刺激に応答して核から移行することが示されている11。コラーゲンへの細胞接着がα1−インテグリンとTCPTPとの同局在化を誘導するので、本発明者らは、マトリックスへの結合またはマイトジェン刺激がその2つのタンパク質の物理的結合を導き得るかどうか調べた。免疫沈降実験において、内因性インテグリンα1β1は、コラーゲンへの接着または血清刺激を受けてHeLa細胞における内因性TCPTPと結合したが、一方ポリ−L−リジン上に置かれた、または血清不含条件に維持された細胞においては、相互作用は検出されなかった(図1C)。さらに、TCPTPは、これらの細胞において血清刺激によってさえフィブロネクチン結合インテグリンα5およびα6と結合できなかった(図1C)。逆免疫沈降実験において、TCPTPは容易にインテグリンα1β1に結合したが、コラーゲンへの接着または血清誘導によってのみ反復して特異的であった(図1D)。本発明者らは、内因性α1インテグリンおよびTCPTPは、ヒト細胞においてコラーゲンまたはマイトジェン刺激に応答して物理的に結合するようになるという結論に達し、原形質膜へのTCPTP移行についての分子的説明を提供する。
【0037】
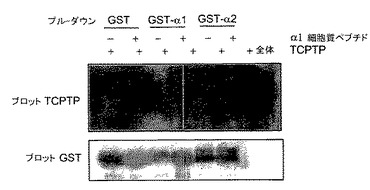
HeLa細胞は高いレベルでα1β1表面発現をし(225±23 FACS平均蛍光)、有意に低いレベルでα2β1インテグリンを発現し(58.5±10.4 FACS平均蛍光)、そしてRT−PCRにより、α10またはα11インテグリンは検出されなかった。Hela細胞において発現されるTCPTPとコラーゲン結合インテグリンとの間の相互作用の特異性を研究するために、本発明者らはプルダウン実験を行った。重要なことには、HeLaライセート由来のGST−融合α1インテグリン細胞質テイル(GST−cytα1)およびGST−融合α2インテグリン細胞質テイル(GST−cytα2)とのプルダウンでは、GST−cytα1のみが内因性TCPTPと結合することがわかった(図1E)。さらに、その相互作用は、精製、組み換えTCPTPがGST−cytα1とは結合するが、GST−cytα2とは結合しないので直接的である(図1F)。プルダウン実験はさらに、すべてのインテグリンαサブユニットと共有する保存されたTrp−Lys−Ile−Gly−Phe−Phe配列(配列番号9)を欠いた可溶性合成ペプチドがTCPTPとの結合においてα1cytと競合することができるので、α1cytとTCPTPとの間の相互作用がα1cytにおける非保存アミノ酸と関係しているということを示した(図1F)。これらのデータは、TCPTPおよびインテグリンの間の相互作用が直接的でありα1β1インテグリンに特異的であることを示す。
【0038】
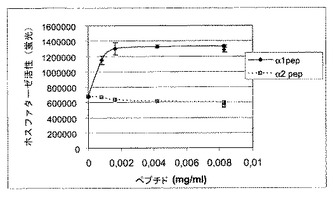
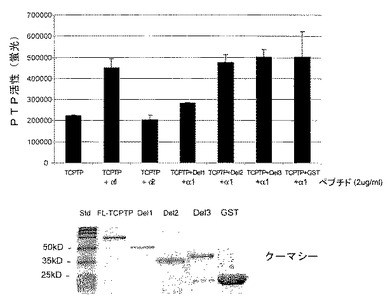
過剰発現試験は、TCPTPが、マイトジェン−およびサイトカイン−誘導シグナル伝達を調節するEGER11、インスリン受容体16およびヤーヌスキナーゼ(JAKs)17などのいくつかの原形質膜結合基質を有するということを示している。インビトロ研究は、TCPTPのタンパク質分解性切断フラグメントを使用し、TCPTPの触媒活性がTCPTPの45kDa型のカルボキシ末端セグメントに関する細胞内阻害によって調節されるということを提案している18。しかしながら、そのような調節機序が細胞内で機能するかどうか、およびそれがどのように作用するかは不明瞭なままである。α1cytおよびTCPTPのカルボキシ末端セグメントは共に同様の塩基性荷電を共有するので、本発明者らは、TCPTPのα1cytとの結合は競合によりこの自動阻害を緩和し、ホスファターゼの活性化を導くという仮説を立てた。実際、コラーゲンへの細胞接着は、特に発現されたTCPTPの触媒活性をポリ−L−リジンに接着した細胞と比較して2.4倍誘導した(図1G)。さらに、a1の細胞質テイルの配列を有する合成ペプチドによる処置に続いて、免疫沈降したTCPTPの触媒活性は2.95±0.48倍に増加した(図2A)。一方、対応するα2テイルペプチドは効果がなかった。活性化は特異的で、他のタンパク質チロシンホスファターゼ、SHP−2はいずれのペプチドによっても活性化されなかった。組み換え、精製TCPTPは、非常に低量の合成α1テイルペプチドにより、高度に活性化された(図2B)。TCPTPのα1cyt誘導活性化の分子的基礎をより詳しく調査するために、組み換え、精製GST−TCPTP欠失変異体(図2C)を、全長TCPTPのα1細胞質ペプチド誘導活性化を妨げるその能力について調べた(図2D)。TCPTPのアミノ末端側半分を含む欠失変異体1が、その活性化を効果的に妨げた。対して、ほとんどのアミノ末端部分が含まれる場合でさえ、またはアミノ末端側半分が欠失されている場合、競合は観察されなかった。したがって、インテグリンα1細胞質テイルはTCPTPのアミノ末端部分と結合し、それを活性化するか、あるいは提案された自動制御カルボキシ末端セグメント18がタンパク質のN−末端部分と相互作用するのを阻害する可能性がある。
【0039】
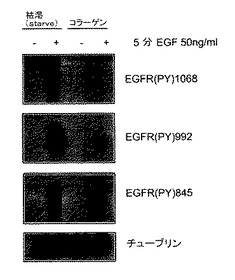
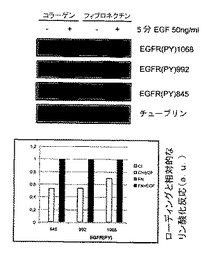
インテグリンと成長因子の媒介シグナルは重なり合い、さらに相乗作用を与える2。マトリックスへの細胞接着は、特に受容体チロシンキナーゼを通した有効なシグナル伝達を可能にする許容性シグナルを伝達する。さらに、インテグリンはEGFRと結合し、そのリン酸化を誘導する。EGFRはまた、TCPTPに対する基質でもあり、ホスファターゼの過剰発現がEGFRのリン酸化とシグナル伝達を抑制することが示されている19。インテグリンα1β1−媒介細胞接着が、インビボで実際にTCPTPを活性化するならば、EGFRが影響を受け得る。したがって、本発明者らはEGFRリン酸化に対するマトリックスへの細胞接着の効果を調べた。プラスチック上に単層に維持するかまたはコラーゲン上に1時間置いた、血清を枯渇させたHeLa細胞を、EGFで5分間処理し、EGFRリン酸化について分析した。EGFは、プラスチック上に維持した細胞において試験したチロシン残基全てにおいて効率的にEGFRリン酸化を誘導した。対してコラーゲン上では、EGF−誘導受容体リン酸化は強く阻害された(図3A)。コラーゲンへの接着は、最も効果的にEGFRリン酸化の初期EGF−誘導ピークを弱めた(図3B)。効果がインテグリン−コラーゲン相互作用により特異的であり、剥離とマトリックスへの再植え付けに関係する一般的な現象ではないということを確認するために、コラーゲンかフィブロネクチンのいずれかの上に置かれたHeLa細胞にEGF刺激を行った。EGF−誘導受容体リン酸化はコラーゲン上よりもフィブロネクチン上の細胞において特により高く(図3C)、コラーゲン結合インテグリンがEGFシグナル伝達の阻害に特に関係しているということを示唆している。
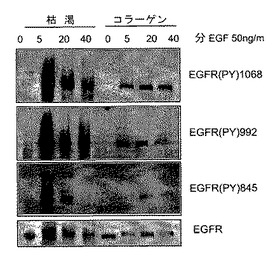
【0040】
本発明者らは、ついでEGFRリン酸化のコラーゲン−誘導阻害がTCPTPの活性化を通して媒介されているかどうかの試験を決定した。TCPTPに特異的な低分子干渉RNAs(siRNA)は、HeLa細胞においてTCPTPの内因性発現を効率的に減少させたが、スクランブル対照siRNAは減少させなかった(図3D)。それらはまた、コラーゲン上に置かれた細胞でEGF−誘導EGFRリン酸化を有意に増加させた(図3D)。これらのノックダウン実験は、EGFRシグナル伝達のコラーゲン誘導阻害におけるTCPTPの役割を確認する。
【0041】
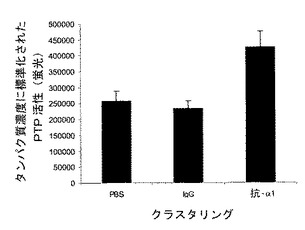
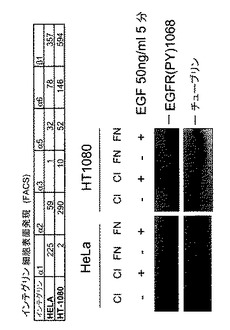
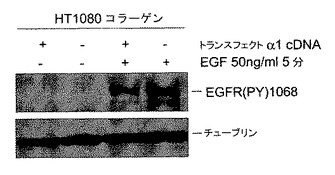
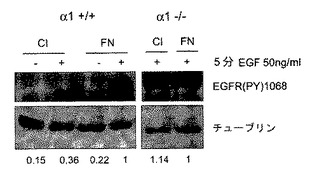
HeLa細胞は、α2β1インテグリンと同様にα1β1インテグリンによりコラーゲンに接着できる。インテグリン−TCPTP−EGFRシグナル伝達経路のインテグリン特異性を試験するために、本発明者らは3つのストラテジーを用いた。第1は、α1β1インテグリンを発現しないHT1080線維肉腫細胞において、コラーゲンへの接着はEGFRリン酸化を阻害することはできなかった(図2E)。重要なことは、一時的にα1インテグリンを発現するようにトランスフェクトされたHT1080細胞は、EGFRリン酸化のコラーゲン誘導阻害に感受性となった(図2F)。第2は、接着、血清枯渇HeLa細胞において、モノクローナル抗体とインテグリンα1−サブユニットのクラスタリングは、細胞質PTP活性を1.5倍増加させ、EGFRのEGF−誘導リン酸化を阻止した(図2G)。最終的に、本発明者らは、α1インテグリンヌルマウスおよびその野生型同腹子由来のマウス胚線維芽細胞を用いてその特異性を調べた7。野生型とα1−/−細胞は両方コラーゲンに接着した9(図3E)。しかしながら、α1β1インテグリンの非存在下では、TCPTPは接着部位に見出されず、一方、野生型細胞ではインテグリンα1β1とTCPTPが細胞の周囲の接着部位に同局在化した(図3E)。α1−/−細胞は、コラーゲンまたはフィブロネクチン上に置かれた場合、同等のEGF−誘導EGFRリン酸化を示し、一方、野生型細胞では、コラーゲンへの接着がEGF−誘導シグナル伝達を阻害する結果となった(図3F)。まとめると、これらの実験はα1−インテグリンがTCPTPを特異的に活性化し、この相互作用がEGFRのリン酸化の阻害をもたらすということを確認する。
【0042】
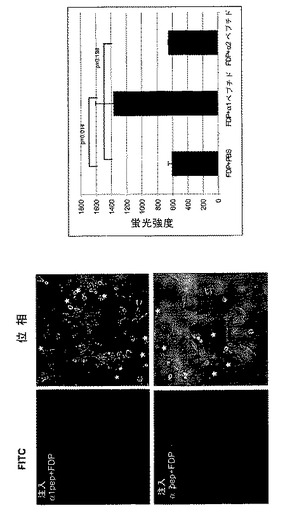
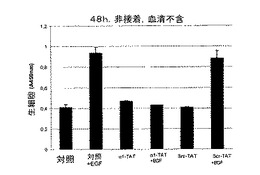
TCPTPの変異体または野生型を用いる過剰発現研究は、細胞増殖の調節におけるその役割を証明した20〜23。さらに、TCPTPは、変異体EGFRを発現するグリア芽腫細胞の増殖および足場非依存性成長を抑制することが示されている23。しかしながら、これまでのところ、TCPTP活性の調節、したがって、その阻害機能のインビボでの実行の機序は知られていない。本発明者らは、インテグリンα1細胞質テイルがインビトロでTCPTPを活性化する能力があることを本明細書に示した。それが細胞においてもホスファターゼ活性を誘導するかどうかを試験するために、本発明者らは、非蛍光ジホスホフルオレセイン(FDP)を、α1またはα2細胞質テイルペプチドと一緒にHeLa細胞中に微量注入した。FDPは脱リン酸化の際に蛍光を発するようになるので、本発明者らは、生HeLa細胞においてホスファターゼ活性を調べた。これらの定量アッセイは、α2またはPBSを同時注入した細胞と比較した場合、インテグリンα1細胞質ペプチドがHeLa細胞においてホスファターゼ活性を誘導したことをはっきりと示した(図4A)。
【0043】
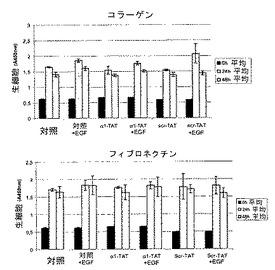
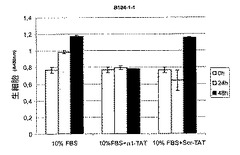
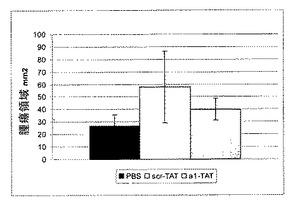
ホスファターゼ活性の誘導は、TCPTPの過剰発現と同様の様式で細胞の腫瘍形成性に影響を与え得る23。最近、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインが、生細胞中にペプチドおよびタンパク質を輸送するために使用されている24、25。したがって、本発明者らは、増殖および腫瘍形成アッセイ用に細胞にα1−cytペプチドを送達するために、TAT−タンパク質に融合した細胞透過性FITC−共役ペプチドを使用した。α1cyt−TATペプチドおよびスクランブル対照ペプチドは両方細胞に効率的に入り(図4B)、蛍光は24時間以上持続し、細胞の剥離は観察されなかった(示さず)。懸濁液中、5%血清で増殖するHeLa細胞では、200nM α1−TATペプチドが血清誘導細胞増殖を阻害した(図4B)。これに対して、対照ペプチドでは増殖は有意に変化しなかった。さらに、α1−TATペプチドはEGF誘導細胞増殖の阻害に有用であるが、懸濁液中の血清不含条件に維持されているHeLa細胞の基礎増殖および生存には効果はなかった(図4D)。腫瘍形成性B104−1−1細胞(neu−腫瘍遺伝子で形質転換された3T3線維芽細胞)の血清誘導増殖もα1−TATペプチドによって阻害された(図4C)。興味深いことに、接着HeLa細胞の増殖は影響されなかったので、α1−TATペプチドは、足場非依存性、EGF−促進(driven)増殖の調節にのみ有効であった(図3G)。さらに、9日間軟寒天で培養されたHeLa細胞による大きな腫瘍コロニーの形成は、α1−TATペプチドによって有意に阻害されたが、対照ペプチドはコロニーサイズを有意に減少させることはできなかった(図4D)。最終的に、インビボでのマウスにおける皮下腫瘍としてのヒト線維肉腫細胞の増殖は、α1−TATペプチドによる処置により阻害された(図5A)。興味深いことに、詳細な免疫組織化学的研究により、α1−TATペプチドで処置した腫瘍は、Scr−TATのPBS注射のいずれかで処置した対照腫瘍よりもより壊死を示すということが明らかとなった(図5B)。これらの実験により、インビボでα1細胞質テイルペプチドによるホスファターゼ活性の誘導が、腫瘍形成性細胞におけるTCPTPの過剰発現により達成される効果と同一の効果をもたらすということが証明される20、23。さらに、ペプチドは生細胞に導入することができ、効率的に悪性細胞の足場非依存性、EGF誘導増殖を阻止し、インビボでの正常細胞増殖の維持におけるα1−送達負シグナルの可能な役割を示唆する。
【0044】
インテグリンのα−細胞質ドメインは、インテグリン媒介生物応答の調節に不可欠であるが26、これまでのところ、ほんの少しの相互作用タンパク質が同定されているのみである。インテグリンは受容体チロシンキナーゼシグナル伝達を正に調節すること、さらにリガンドの非存在下でRTK活性化を誘発することが知られている3、27。また、遍在して発現されるインテグリンα1は、コラーゲンに応答して一次細胞でERK MAP−キナーゼ経路を活性化することが示されている9。他方では、インテグリンα1は、乳癌、卵巣癌および肺腺癌においてダウンレギュレートされると思われる3、27、28。TCPTPのインテグリンα1β1媒介活性化によるEGFRシグナル伝達の負の調節は、細胞応答の接着媒介制御における新規な機序であると思われる。これらの知見は、細胞におけるこの腫瘍抑制タンパク質のホスファターゼ活性の制御に対する分子的説明を提供し、なぜTCPTP(他のPTPsのように)が単離タンパク質としては不規則であるのに、細胞においては高度に特異的であるのかという1つの機序を証明する29。インテグリンα1によるTCPTPの局所調節はそのうえ、タンパク質リン酸化−脱リン酸化平衡の正確な局在化制御が不可欠である他のシグナル伝達経路にとっても重要なようである。たとえば、(インテグリンと結合する27)EGFRと極めて接近した原形質膜でのインテグリンα1細胞質ドメインとの相互作用によるTCPTPの局在的活性化は、厳重に調節された方法で、RTKシグナル伝達を制御するための細胞にとっての強力な方法である。α1細胞質ペプチドの生細胞への送達による、細胞の、接着ではなく足場非依存性増殖の阻害は、形質転換細胞を標的化する新しい手段を提供し得る。結論として、腫瘍抑制タンパク質のインテグリンα1β1媒介活性化によるEGFRのシグナル伝達の負の調節は、細胞応答の接着媒介制御における新規なパラダイムである。
【0045】
本発明の方法は、さまざまな実施態様の形態で組み込まれ得、そのうちのほんのいくつかを本明細書に開示していることは認識されよう。当業者には、他の実施態様が存在し、それは本発明の精神から逸脱しないことが自明である。したがって、記載した実施形態は例示であり、制限的なものと解釈されるべきでない。
【0046】

【図面の簡単な説明】
【0047】
【図1A】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。PC3細胞を、1時間コラーゲン、フィブロネクチンまたはポリ−L−リジンに接着した。インテグリンα1サブユニットおよびTCPTPが二色免疫蛍光染色により検出された。矢印は、膜での同局在化の各領域をそれぞれ指し示す。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して評価した、バー5μm。
【図1B】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞を、1時間コラーゲン、フィブロネクチンまたはポリ−L−リジンに接着した。インテグリンα1サブユニットおよびTCPTPが二色免疫蛍光染色により検出された。矢印は、膜での同局在化の各領域をそれぞれ指し示す。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して評価した、バー5μm。
【図1C】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。血清枯渇HeLa細胞を表面ビオチン化し、プラスチック上に置き、10%FBSで30分間刺激する(FBS)か、コラーゲンI(CI)またはポリ−L−リジン(PL)上に1時間置いた。免疫沈降(ip)されたインテグリン、TCPTPおよび対照(IgG)を免疫ブロッディングで検出した。
【図1D】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞を図1Cと同様に処理し、ライセートを免疫沈降し(ip)、TCPTPに対しブロットするか、または変性させ、抗−α1抗体で再沈降(re−ip)させ、示したように免疫ブロットした。
【図1E】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞ライセートを固定化グルタチオン−S−トランスフェラーゼ(GST)またはGST−インテグリン細胞質ドメイン融合タンパク質と共にインキュベートした。結合したタンパク質およびライセート試料をTCPTPについてプローブした。ポンソーS染色をローディング対照に使用した。
【図1F】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。ポンソーS染色をローディング対照に使用した。組み換えおよび精製TCPTP(1μg)をGSTまたはGST−融合タンパク質と、α1インテグリン細胞質ドメインペプチド(1μg/ml)と共にまたはなしでインキュベートした。結合したタンパク質(およびTCPTPローディング対照、全体)をTCPTPおよびGSTについてプローブした。
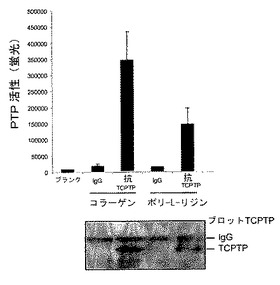
【図1G】血清枯渇HeLa細胞を剥離し、コラーゲンまたはポリ−L−リジン上に置き、免疫沈降(ip)にかけ、そしてホスファターゼ活性(平均±SD、n=3)を測定した。試料の半分をTCPTPについてイムノブロットした。
【図1H】血清枯渇HeLa細胞をPBS、対照IgG、または抗−α1 mAbとインキュベートし、抗−マウス二次抗体により30分間クラスタリングを誘導した。ライセート由来の等量のタンパク質のホスファターゼ活性(平均±SD、n=3)を三重で分析した。
【図2A】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。HeLa細胞を、対照(IgG)、抗−TCPTPおよび抗−SHP−2抗体と共に免疫沈降(ip)した。ビヒクル(c)および合成α1細胞質テイルペプチド(α1pep)および合成α2細胞質テイルペプチド(α2pep)での処置後、ホスファターゼ活性(平均±SD、n=3)を、diFMUPを基質として用いて分析した。免疫沈殿物の半分をSDS−PAGE上で分離し、TCPTPまたはSHP−2について標識した。
【図2B】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。組み換え、精製TCPTP(0.15μg/ml)を、異なる濃度のペプチドとインキュベートし、ホスファターゼ活性(平均±SD、n=3)を分析した。
【図2C】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。TCPTP欠失変異体の概略的ダイアグラムを示す。
【図2D】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。競合アッセイでは、全長TCPTPを添加する前に、GSTと融合したTCPTP欠失変異体、またはGSTのみを示したペプチドと共にインキュベートし、ホスファターゼ活性(平均±SD、n=3)を測定した。
【図2E】インテグリンα1β1がEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。FACSによってインテグリンの表面発現をHeLa細胞およびHT1080細胞で分析した。MFI値を示す。血清枯渇HeLa細胞およびHT1080細胞をコラーゲンI(CI)またはフィブロネクチン(FN)上に置き、EGFで処置した。EGFRリン酸化をホスホ特異的抗体を用いて調べた。チューブリンをローディング対照として使用した。
【図2F】インテグリンα1β1はEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。α1−cDNAで一時的にトランスフェクトされたHT1080細胞またはモック−トランスフェクトHT1080細胞から血清を枯渇させ、コラーゲンI(CI)上に置き、EGFで処理した。EGFRリン酸化は図2Eと同様にして分析した。
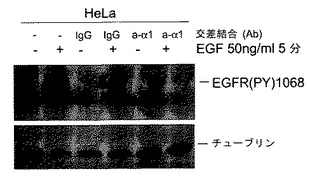
【図2G】インテグリンα1β1はEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。血清枯渇HeLa細胞を図1Hと同様に処理してα1受容体と交差させ、EGFRリン酸化をイムノブロッディングにより調べた。
【図3A】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持された、またはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。チューブリンをローディング対照として使用した。
【図3B】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持されたまたはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。EGFRをローディング対照として使用した。
【図3C】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持されたまたはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。3つ(A、C)の実験のデンシトメトリーによる分析(平均±SD)が示される。チューブリンをローディング対照として使用した。
【図3D】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。TCPTPに対して特異的な2つのsiRNAs(またはスクランブル対照)でトランスフェクトされたHeLa細胞を、コラーゲン上に置き、EGFで処理した。抽出物を示したタンパク質についてイムノブロットした。同様の結果を有する3つの実験の代表を示す。
【図3E】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。α1−/−マウスまたはその野生型同腹子由来の線維芽細胞をコラーゲン上に置き、インテグリンα1およびTCPTPに対して免疫染色した。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して分析した;バー5μm。
【図3F】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を軽減することを示す。α1−/−および+/+動物由来の血清枯渇線維芽細胞をコラーゲンまたはフィブロネクチン上に置き、EGFで処理し、イムノブロットした。値はチューブリンに標準化したデンシトメトリーによる評価法である。
【図3G】インテグリンα1細胞質ペプチドがマトリックス接着細胞のEGF非依存性増殖に影響を及ぼさないことを示す。HeLa細胞をコラーゲンまたはフィブロネクチンでコートしたウェル中、5%血清で、200nM TATペプチドおよび50ng/ml EGFの存在下、または非存在下で培養し、種々の時点での生細胞の数(平均±SD、n=3)を分析した。
【図4A】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。HeLa細胞を、脱リン酸化の際に蛍光を発するようになるフルオレセインジホスフェート(FDP)と細胞質インテグリンペプチドと共に微量注入した(アスタリスク)。蛍光強度を30分間モニターし(30分での代表的な画像を示す。)、個々の細胞における平均強度(平均±SEM、n=12)を評価した。
【図4B】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。FITC標識TAT−α1細胞質テイル融合ペプチド(α1−TAT)およびTAT−スクランブル対照融合ペプチド(Scr−TAT)をHeLa細胞に入れたことを示す。5%血清、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での非接着生HeLa細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4C】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。血清不含培地で、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での非接着生HeLa細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4D】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。血清含有培地で、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での腫瘍形成性B104−1−1線維芽細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4E】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。HeLa細胞を寒天培地中でTAT−ペプチドと共に、またはなしで9日間増殖させた。代表的な位相差画像を二重盲式で撮影し、コロニーの大きさの分析を示す。
【図5A】α1細胞質テイルペプチドが腫瘍増殖を減少させ、インビボで壊死を誘導することを示す。HT1080ヒト線維肉腫細胞をヌードマウスの側面に注射し、実験項に示したように処理した。皮下腫瘍は4週間後に取り出した。腫瘍の大きさ(領域)を測り、示す(平均±SEM、n=12)。
【図5B】α1細胞質テイルペプチドが腫瘍増殖を減少させ、インビボで壊死を誘導することを示す。HT1080ヒト線維肉腫細胞をヌードマウスの側面に注射し、実験項に示したように処理した。皮下腫瘍は4週間後に取り出した。全腫瘍切断面をマッソン(Masson)トリクロム染色で染色し、全腫瘍領域に対する壊死領域を二重盲式で記録した(平均±SEM、n=12)。壊死領域の分析を示す。
【技術分野】
【0001】
本発明は、T細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法および個体におけるチロシンキナーゼシグナル伝達の阻害方法に関する。さらに、本発明は個体におけるチロシンキナーゼシグナル伝達を阻害することにより治癒可能である疾患または障害の予防または治療方法に関する。さらにまた、本発明は、T細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化に基づく、個体における癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法に関する。本発明はまた、前記方法に有用な医薬組成物に関する。
【背景技術】
【0002】
本発明の背景を説明するために本明細書で使用する刊行物および他の資料、および特に、実施に関するさらなる詳細を提供するための事例は、引用により本明細書に組み込まれる。
【0003】
インテグリン媒介細胞接着は、増殖、生存および異なる細胞性シグナル伝達経路間のクロストークなどの多様な細胞応答を制御する1。これまでのところ、インテグリンは、足場依存性受容体チロシンキナーゼシグナル伝達を可能にする許可シグナルを伝えることが主に示されている2、3。細胞接着への応答において、インテグリンはある細胞質シグナル伝達経路を直接活性化し、間接的に成長因子受容体によるシグナル伝達を調節する。インテグリンの細胞質ドメインは、これらの機能を媒介するのに不可欠である4〜7。しかしながら、それらは固有の触媒活性を欠き、シグナル伝達のために細胞質タンパク質との相互作用を必要とする。コラーゲンは人体に非常に豊富に存在するタンパク質であるが、4つのコラーゲン結合インテグリンα1β1、α2β1、α10β1およびα11β1のシグナル伝達については相対的にほとんど知られていない。インテグリンα1β1は、コラーゲンとラミニンの受容体である。その1つのサブユニットは、カベオリン−1と関連していることが示されており、したがって、シグナル伝達アダプタータンパク質Shcを補充する(recruit)8。これは次にマイトジェン活性化タンパク質キナーゼ経路の活性化ならびにコラーゲン上の線維芽細胞の生存および増殖の増強をもたらす9。さらに、すべてのインテグリン細胞質テイルにみられる保存領域は、パキシリン、テーリン(talin)および焦点接着キナーゼとのつながりによって、焦点接着集合における機能が示されている10。しかしながら、これまでのところα1インテグリンにより明確に活性化されるシグナル伝達経路は確認されていない。
【発明の開示】
【0004】
本発明は、α−1−インテグリンの細胞質テイルが選択的かつ直接的に遍在的に発現するタンパク質チロシンホスファターゼTCPTP(T細胞タンパク質チロシンホスファターゼ)と相互作用し、コラーゲンへの細胞接着の際にそれを活性化させるという発見に基づく。その活性化により、EGF刺激の際のEGFR(上皮成長因子受容体)リン酸化が減少することとなる。α−1−インテグリンの細胞質テイルは、腫瘍抑制タンパク質、TCPTPの活性化を通してEGFRシグナル伝達の負の調節因子として機能する。α−1−細胞質ドメインペプチドの細胞への導入により、ホスファターゼ活性が誘導され、インビボでのヒト線維肉腫の接ぎ木(zenografts)においてはもとより、EGF誘導細胞増殖およびインビトロにおける悪性腫瘍の足場非依存性成長を誘導する。これらのデータは、インビボにおけるTCPTP活性の制御の最初の証明であり、受容体チロシンキナーゼシグナル伝達のインテグリン媒介性負調節の新規な分子パラダイムを表している。
【0005】
したがって、1つの側面によれば、本発明は、治療に使用するためのi)アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチド、またはii)哺乳類細胞において該ペプチドを発現可能なベクターのいずれかである薬剤に関する。
【0006】
もう1つの側面によれば、本発明は個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法であって、該個体に、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである有効量の薬剤を投与することによる方法に関する。
【0007】
第3の側面によれば、本発明は個体におけるチロシンキナーゼシグナル伝達の阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0008】
第4の側面によれば、本発明は個体における疾患または障害の予防または治療方法であって、該疾患または障害がチロシンキナーゼシグナル伝達を阻害することにより治癒可能なものであり、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0009】
第5の側面によれば、本発明は個体における、癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法に関する。
【0010】
第6の側面によれば、本発明は、治療に効果的な量のTCPTPを活性化できる低分子またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれか、および薬学的に許容され得る担体を含む医薬組成物に関する。
【0011】
第7の側面によれば、本発明は、アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドをコードする核酸を包含する発現ベクターであり、哺乳動物細胞において該ペプチドを発現することができるベクター、および薬学的に許容され得る担体を含む医薬組成物に関する。
【発明を実施するための最良の形態】
【0012】
定義:
用語「治療」または「治療する」は、疾患または障害の完全な治癒ならびに該疾患または障害の改善または軽減を含むことを理解されたい。
【0013】
用語「予防」は、完全な予防(prevention)、防御(prophylaxis)、および前記疾患または状態を伴う病気になる個体のリスクを低下することを含むことを理解されたい。
【0014】
用語「個体」は、ヒトまたは動物の被験体を意味する。
【0015】
用語「有効量」は、特に動物またはヒトの被験体に投与する際に、所望の治療結果をもたらすのに充分な本発明の薬剤の任意の量を意味する。
【0016】
好ましい実施態様:
TCPTPの活性化に使用される薬剤は、TCPTPを活性化できる低分子であっても、またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれであってもよい。この配列は、α−1−インテグリンの細胞質テイルに特異的である。1つの実施態様によれば、ペプチドは正にアミノ酸配列RPLKKKMEK(配列番号1)であってよく、たとえば微量注入によって細胞に投与され得る。
【0017】
あるいは、ペプチドは、アミノ酸配列RPLKKKMEK(配列番号1)を包含するより長い鎖であってもよい。特に好ましい実施態様によれば、ペプチド(鎖RPLKKKMEK(配列番号1)またはそれを包含するより長い鎖)は、細胞膜透過性融合タンパク質の一部であってもよい。そのような融合タンパク質の例としては、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質転換ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む融合タンパク質があげられる。そのような融合タンパク質の例としては、YGRKKRRQRRRWKLGFFKRPLKKKMEK(配列番号2)があげられる。配列YGRKKRRQRRR(配列番号3)は、TATから誘導され、配列WKLGFFK(配列番号4)は、インテグリン(いずれのαインテグリンにも典型的な)から誘導される。
【0018】
さらなる代替によれば、投与される薬剤は、哺乳動物細胞において所望のペプチドを発現することができる核酸を含むベクターであってもよい。核酸はDNA配列中、RNA配列中またはウイルスベクター中に挿入することができる。そのようなウイルスベクターは、通常、アデノウイルス、アルファウイルス、アデノ随伴ウイルス、レトロウイルスまたはヘルペスウイルスに基づく。細胞におけるペプチドの発現については、たとえばM Parsonsら、Molecular and Cellular Biology, Aug. 2002, p.5897-5911.参照。
【0019】
よって、本発明によるTCPTPの活性化は、直接的または間接的に該活性化によって影響される障害または疾患を予防または治療するために使用され得る。
【0020】
少なくとも部分的に間接的に影響される疾患または障害の例としては、その治癒性がチロシンキナーゼシグナル伝達を阻害することにより恩恵を受け得る任意の疾患または障害があげられる。具体的には、そのような標的の非限定的な例は、上皮細胞成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼがあげられる。EGFRの阻害は、癌の予防、癌の成長、浸潤もしくは転移の予防または阻害に有用であり得る。しかしながら、これらの効果も、TCPTPの活性化による他の機序により部分的となり得るかもしれない。
【0021】
他方、EGFRの阻害はまた、他の、非癌疾患または障害の予防または障害に有用であり得る。
【0022】
インスリン受容体およびヤーヌスキナーゼの阻害は、インスリン受容体によって制御されるエネルギー代謝の異常を誘導するインスリン初期シグナル伝達の亢進と関係する疾患、ならびにインターフェロン受容体と結合するヤーヌスキナーゼの異常な活性を導くインターフェロンによる望ましくない炎症性シグナル伝達と関係する障害の治療または予防に有用であり得る。
【0023】
配列RPLKKKMEK(配列1)を含むペプチドは、細胞膜を通して透過できる形態でもたらされるのが好ましい。そのようなペプチドは、組成物の非経口投与に適した任意の担体と投与することができる。たとえば、ペプチドは脂質と複合化されるか、リポソームに詰められるか、シクロデキストリンもしくは他の複合化剤、生吸収性(bioresorbable)ポリマーまたは制御された放出投与のための他の好適な担体、ナノ粒子またはヒドロゲルに組み込まれることができる。
【0024】
あるいは、アミノ酸配列RPLKKKMEK(配列1)を含むペプチドをコードする核酸を包含する発現ベクターを個体に投与することもできる。核酸は、正に配列RPLKKKMEK(配列1)、またはこの配列を含むより長い配列をコードすることができる。ベクターは、たとえば脂質と複合化されるか、リポソームに詰められるか、シクロデキストリンもしくは他の複合化剤、生吸収性ポリマーまたは制御された放出投与のための他の好適な担体、ナノ粒子またはヒドロゲルに組み込まれることができる。
【0025】
そのような治療を必要とする患者に投与されるペプチドまたは発現ベクターの治療的有効量は、たとえば患者の年齢および体重、治療を必要とする正確な病状、およびその重篤度、および投与経路などの多数の因子に依存し得る。正確な量は、最終的には主治医の裁量であろう。したがって、本発明の実施は任意の用量、他の治療有効薬との組み合わせ、非経口投与のための薬学製剤またはデリバリーシステムを含む。ペプチドまたは発現ベクターは、全身的または局所的に投与される。適切な投与経路としては、静脈内、筋肉内、皮下注射、吸入、局所、眼、舌下、鼻、直腸、腹腔内デリバリー、およびイオン導入または他の経皮デリバリーシステムがあげられる。
【0026】
本発明を、以下の非制限的な実施例の項により説明する。
【実施例】
【0027】
方法
酵母ツーハイブリッドシステム
α1−インテグリン細胞質ドメインを、pGBKT7ベクター(クローンテック)におけるGAL4 DNA−BDに融合し、マウス17日胚適合マーカーcDNAライブラリー(BDクローンテック)をスクリーニングするためのベイトとして使用した。そのライブラリーのスクリーニングは、製造元のプロトコールにしたがって行なった。単離した独立したクローンの1つがGAL4−DBD−α1cytと相互作用したが、T細胞タンパク質チロシンホスファターゼをコードする無関係なベイトとは相互作用しなかった。
【0028】
タンパク質−タンパク質相互作用アッセイ
α1cytおよびα2cytをpGEX4T1(アマシャムバイオサイエンス)にサブクローニングした。全長TCPTP(45kD)およびその欠失変異体をpCGTC45プラスミド11からPCR増幅し、pGEX−6P−1にクローニングした。GST融合タンパク質をE.coli(BL21 pLysS)で発現させ、製造元(アマシャムバイオサイエンス)の取扱説明書にしたがい精製した。融合タンパク質をグルタチオンセファロースに固定し、25mMグルタチオンで溶出するか、またはGSTを製造元の取扱説明書にしたがって正確な(precession)プロテアーゼを用いて切断した。プルダウンアッセイについては、Hela細胞を溶解緩衝液(1%オクチルグルコシド、50mMトリス−HCl pH7.5、150mM NaCl、1mM MgCl2、完全プロテアーゼ阻害剤(ロッシュ))に溶解させ、遠心分離、10分、13000rpm+4Cで浄化(clarified)し、グルタチオンセファロースで前除去し(precleared)、そして固定化GST融合タンパク質とインキュベートし、同じ緩衝液1mlで3回洗浄した。結合したタンパク質をSDS−PAGEで分離し、ウエスタンブロットで検出した。あるいは、精製全長TCPTPを溶解緩衝液中で、合成インテグリンα1細胞質テイルペプチド(RPLKKKMEKPLKKKMEK(配列番号5)ジェネムド シンセシス)の存在下または非存在下で示すようにGST−融合タンパク質とインキュベートした。
【0029】
免疫沈降、ウエスタンブロッディングおよびホスファターゼアッセイ
血清枯渇(starved)Hela細胞をプラスチック上に未処理のまま置き、10%FBSで30分刺激するか、コラーゲンI型またはポリ−L−リジン(10μg/ml、シグマ)で被覆した組織培養プレート上に1時間置いた。示した場合、細胞は記述したように表面をビオチン化した12。細胞をLaemmli試料緩衝液に溶解させウエスタンブロット用にSDS−PAGE上で分画するか、または溶解緩衝液に溶解させ、各試料由来のタンパク質をそれぞれ等量Gタンパク質−セファロースで前除去し免疫沈降に付した。抗体(抗−TCPTP、オンコジーン;抗−a1、抗−a5、抗−a6、すべてケミコン;抗−SHP−2、サンタクルーズ)をGタンパク質−セファロースと共にインキュベートし、免疫沈降を4℃で2時間行なった。免疫沈殿物を細胞溶解緩衝液で3回洗浄した。共沈殿アッセイについては、タンパク質をSDS−PAGEで分離し、ウエスタンブロットで検出するか、Vectastain試薬(ベクター)でビオチンの検出を行なった。ホスファターゼアッセイについては、免疫沈殿物をホスファターゼ反応緩衝液(25mM Hepes pH7.4、50mM NaCl、1mM DTT)に再懸濁させ、反応物の1/3をウエスタンブロッディングに付し、残りのビーズを、基質としてdiFMUP(モレキュラープローブス)を用い、セリン/スレオニンホスファターゼ阻害剤カクテル(シグマ)の存在下で製造元の取扱説明書にしたがって三重でホスファターゼ活性を評価した。示した場合、合成ペプチドは、ホスファターゼアッセイ反応の前に15分間免疫沈殿物とプレインキュベートされた(α2ペプチドKLGFFKRKYEKMTKNPDEIDETTELSS(配列番号6)、Heino博士より提供)。精製タンパク質とのホスファターゼアッセイについては、全長TCPTPをホスファターゼ反応緩衝液で、示したようにTCPTP欠失変異タンパク質および合成インテグリン細胞質テイルペプチドの存在下または非存在下でインキュベートした。ウエスタンブロッディングをEGFR、EGFR(PY)845、992もしくは1068(セルシグナリングテクノロジーズ)、TCPTP(オンコジーン)またはチューブリン(サンタクルーズバイオテクノロジー)に対して作製された市販の抗体で行なった。抗体の特異的結合を過酸化水素共役二次抗体で検出し、拡張化学発光検出法により可視化した。
【0030】
トランスフェクション
HT1080細胞を、以前に報告されているように13、Fugene6(ロッシュ)試薬を用いてpcDNA3.1/zeoベクター(バンダービルト大学、Pozzi博士より提供)中のα1cDNAで一時的にトランスフェクトした。TCPTPを標的とする2つの異なるアニールしたsiRNAs(ggcacaaaggaguuacauctt(配列番号7)、ggaguuacaucuuaacacatt(配列番号8);アンビオン(Anbion))またはスクランブル対照siRNAを、Hela細胞にオリゴフェクトアミン(インビトロジェン)を用い、製造元のプロトコールにしたがってトランスフェクトした。24時間後、トランスフェクション細胞を24時間血清枯渇させ、溶解およびウエスタンブロットアッセイの前に示したように処理した。
【0031】
免疫蛍光
I型コラーゲン、フィブロネクチンまたはポリ−L−リジン(10μg/ml)でコートした酸洗浄ガラスカバースリップ上に細胞を置き、1時間接着させた。細胞をPBSで洗浄し、4%パラホルムアルデヒドで10分間固定した。抗体染色については、細胞をPBS/0.1% Triton X−100に浸透させ(permeabilized)、ブロッキングのためにPBS/1%(w/v)BSAで洗浄した。細胞を室温で1時間、示した抗体で染色した。ついで、Alexa−488またはAlexa−555で共役された抗ウサギまたは抗マウス抗体(モレキュラープローブス)による検出の前に、細胞をPBSで3回洗浄した。PBSと水とで洗浄後、細胞をMowiol(100mM Tris−HCl pH8.5、10%(w/v)Mowiol(カルビオケム、サンディエゴ、CA)および25%(v/v)グリセロール含有アンチフェイド(antifade)(2.5%(w/v)1,4−ジアザジシクロ−2.2.2−オクタン(シグマ)))に載せた。スライドは、ツァィス倒立蛍光顕微鏡(Zeiss inverted fluorescence microscope)または共焦点レーザー顕微鏡(Axioplan 2 LSM 510;カールツァィス社(Carl Zeiss Inc.)、イェーナ、ドイツ)(63×/1.4 Plan−Apochromat油浸対物レンズを装備)を用いて調べた。共焦点画像は、約1.0μmの単一z−断面を表す。PBS中、1mM FDP(モレキュラープローブス)および1mg/mlのペプチドによる細胞質微量注入法を、ガラスの底3cmプレート上で培養されるHela細胞に行なった。生細胞の蛍光画像は、同一の設定で撮り、強度はZeiss Axioplan 4ソフトウエアを用いて定量化した。
【0032】
細胞増殖および軟寒天アッセイ
96ウェルプレートをBSAまたは示したマトリックスでコートした。細胞をそのウェルに104/ウェルで播いた。TATペプチド(α1−TAT fitc−YGRKKRRQRRRWKLGFFKRPLKKKMEK(配列番号2)、Scr−TAT fitc−YGRKKRRQRRRLKGWRFKLKPKFKEMK(配列番号2);ジェネムドシンセシス)およびEGF(シグマ)を示したように添加し、生存細胞の数をWST−1(ロッシュ)で製造元のプロトコールにしたがって検出した。軟寒天アッセイは記載されたように行なった14。200nM TATペプチドを含む培地を48時間毎に置き換え、9日間のインキュベーション後、培養物を二重盲検で撮影した。各視野(4×倍率)由来のコロニーの数と大きさをシンジーン(Syngene)のGeneToolsソフトウエアを用いて分析した。コロニーを大きさで分類した(小=200〜499ピクセル、中=500〜1000および大1000以上)。
【0033】
マウスおよび腫瘍細胞播種
雄の無菌ヌードマウスを4週齢で使用した。HT1080腫瘍細胞(5×106細胞/100μl PBS)をヌードマウスの背面右脇腹に皮下注射した。マウスを腫瘍細胞播種部位に隣接した皮下注射(20μl PBS、PBS中の20μM a1−TATまたは20μM Scr−TATペプチド)で、実験期間中週に3回処置した。播種後4週間でマウスを犠牲死させた。腫瘍を取り出し、その大きさを定量し、免疫組織化学用にパラフィンブロック中に加工した。IHCを以前に記載されたように行なった(Grant D.S.ら、Int. J. Cancer 2003 104: 121-129)。
【0034】
結果:
コラーゲンへのα1インテグリン−媒介細胞接着によって活性化されるシグナル伝達経路を特徴付けるために、本発明者らは、α1細胞質ドメイン(α1cyt)と相互作用するタンパク質を同定するために酵母ツーハイブリッドシステムを使用した。陽性クローンの1つは、TCPTPの45kDa型をコードしていた15。この分子は、遍在的に発現される核非受容体タンパク質チロシンホスファターゼであり、マイトジェンの刺激に応答して細胞質に移行することができる11。
【0035】
ヒト細胞における新規な相互作用を実証するために、本発明者らは、まず内因性α1インテグリンとTCPTPとが同じ場所に局在化する(colocalize)かどうかを調べた。PC3およびHeLa細胞を種々のマトリックスタンパク質上に置き、a1−インテグリンおよびTCPTPを二色免疫蛍光染色により可視化した。インテグリンを活性化できないポリ−L−リジン上に置かれた細胞では、TCPTPは細胞質および核の両方で見られた。4つのコラーゲン結合インテグリン以外のインテグリンに結合するフィブロネクチン上に置かれた細胞では、TCPTPは細胞質ゾルに広く見られた(図1B)。対照的に、コラーゲンへの結合では、TCPTPは、細胞膜の末梢領域においてα1β1インテグリンと同じ場所に局在化した(図1A、B)。これらのデータは、内因性インテグリンα1とTCPTPとがコラーゲンへの細胞接着により特異的にヒト細胞において同局在化することを示す。
【0036】
以前の研究では、TCPTPが分子的には特徴付けされていない機序によりマイトジェン刺激に応答して核から移行することが示されている11。コラーゲンへの細胞接着がα1−インテグリンとTCPTPとの同局在化を誘導するので、本発明者らは、マトリックスへの結合またはマイトジェン刺激がその2つのタンパク質の物理的結合を導き得るかどうか調べた。免疫沈降実験において、内因性インテグリンα1β1は、コラーゲンへの接着または血清刺激を受けてHeLa細胞における内因性TCPTPと結合したが、一方ポリ−L−リジン上に置かれた、または血清不含条件に維持された細胞においては、相互作用は検出されなかった(図1C)。さらに、TCPTPは、これらの細胞において血清刺激によってさえフィブロネクチン結合インテグリンα5およびα6と結合できなかった(図1C)。逆免疫沈降実験において、TCPTPは容易にインテグリンα1β1に結合したが、コラーゲンへの接着または血清誘導によってのみ反復して特異的であった(図1D)。本発明者らは、内因性α1インテグリンおよびTCPTPは、ヒト細胞においてコラーゲンまたはマイトジェン刺激に応答して物理的に結合するようになるという結論に達し、原形質膜へのTCPTP移行についての分子的説明を提供する。
【0037】
HeLa細胞は高いレベルでα1β1表面発現をし(225±23 FACS平均蛍光)、有意に低いレベルでα2β1インテグリンを発現し(58.5±10.4 FACS平均蛍光)、そしてRT−PCRにより、α10またはα11インテグリンは検出されなかった。Hela細胞において発現されるTCPTPとコラーゲン結合インテグリンとの間の相互作用の特異性を研究するために、本発明者らはプルダウン実験を行った。重要なことには、HeLaライセート由来のGST−融合α1インテグリン細胞質テイル(GST−cytα1)およびGST−融合α2インテグリン細胞質テイル(GST−cytα2)とのプルダウンでは、GST−cytα1のみが内因性TCPTPと結合することがわかった(図1E)。さらに、その相互作用は、精製、組み換えTCPTPがGST−cytα1とは結合するが、GST−cytα2とは結合しないので直接的である(図1F)。プルダウン実験はさらに、すべてのインテグリンαサブユニットと共有する保存されたTrp−Lys−Ile−Gly−Phe−Phe配列(配列番号9)を欠いた可溶性合成ペプチドがTCPTPとの結合においてα1cytと競合することができるので、α1cytとTCPTPとの間の相互作用がα1cytにおける非保存アミノ酸と関係しているということを示した(図1F)。これらのデータは、TCPTPおよびインテグリンの間の相互作用が直接的でありα1β1インテグリンに特異的であることを示す。
【0038】
過剰発現試験は、TCPTPが、マイトジェン−およびサイトカイン−誘導シグナル伝達を調節するEGER11、インスリン受容体16およびヤーヌスキナーゼ(JAKs)17などのいくつかの原形質膜結合基質を有するということを示している。インビトロ研究は、TCPTPのタンパク質分解性切断フラグメントを使用し、TCPTPの触媒活性がTCPTPの45kDa型のカルボキシ末端セグメントに関する細胞内阻害によって調節されるということを提案している18。しかしながら、そのような調節機序が細胞内で機能するかどうか、およびそれがどのように作用するかは不明瞭なままである。α1cytおよびTCPTPのカルボキシ末端セグメントは共に同様の塩基性荷電を共有するので、本発明者らは、TCPTPのα1cytとの結合は競合によりこの自動阻害を緩和し、ホスファターゼの活性化を導くという仮説を立てた。実際、コラーゲンへの細胞接着は、特に発現されたTCPTPの触媒活性をポリ−L−リジンに接着した細胞と比較して2.4倍誘導した(図1G)。さらに、a1の細胞質テイルの配列を有する合成ペプチドによる処置に続いて、免疫沈降したTCPTPの触媒活性は2.95±0.48倍に増加した(図2A)。一方、対応するα2テイルペプチドは効果がなかった。活性化は特異的で、他のタンパク質チロシンホスファターゼ、SHP−2はいずれのペプチドによっても活性化されなかった。組み換え、精製TCPTPは、非常に低量の合成α1テイルペプチドにより、高度に活性化された(図2B)。TCPTPのα1cyt誘導活性化の分子的基礎をより詳しく調査するために、組み換え、精製GST−TCPTP欠失変異体(図2C)を、全長TCPTPのα1細胞質ペプチド誘導活性化を妨げるその能力について調べた(図2D)。TCPTPのアミノ末端側半分を含む欠失変異体1が、その活性化を効果的に妨げた。対して、ほとんどのアミノ末端部分が含まれる場合でさえ、またはアミノ末端側半分が欠失されている場合、競合は観察されなかった。したがって、インテグリンα1細胞質テイルはTCPTPのアミノ末端部分と結合し、それを活性化するか、あるいは提案された自動制御カルボキシ末端セグメント18がタンパク質のN−末端部分と相互作用するのを阻害する可能性がある。
【0039】
インテグリンと成長因子の媒介シグナルは重なり合い、さらに相乗作用を与える2。マトリックスへの細胞接着は、特に受容体チロシンキナーゼを通した有効なシグナル伝達を可能にする許容性シグナルを伝達する。さらに、インテグリンはEGFRと結合し、そのリン酸化を誘導する。EGFRはまた、TCPTPに対する基質でもあり、ホスファターゼの過剰発現がEGFRのリン酸化とシグナル伝達を抑制することが示されている19。インテグリンα1β1−媒介細胞接着が、インビボで実際にTCPTPを活性化するならば、EGFRが影響を受け得る。したがって、本発明者らはEGFRリン酸化に対するマトリックスへの細胞接着の効果を調べた。プラスチック上に単層に維持するかまたはコラーゲン上に1時間置いた、血清を枯渇させたHeLa細胞を、EGFで5分間処理し、EGFRリン酸化について分析した。EGFは、プラスチック上に維持した細胞において試験したチロシン残基全てにおいて効率的にEGFRリン酸化を誘導した。対してコラーゲン上では、EGF−誘導受容体リン酸化は強く阻害された(図3A)。コラーゲンへの接着は、最も効果的にEGFRリン酸化の初期EGF−誘導ピークを弱めた(図3B)。効果がインテグリン−コラーゲン相互作用により特異的であり、剥離とマトリックスへの再植え付けに関係する一般的な現象ではないということを確認するために、コラーゲンかフィブロネクチンのいずれかの上に置かれたHeLa細胞にEGF刺激を行った。EGF−誘導受容体リン酸化はコラーゲン上よりもフィブロネクチン上の細胞において特により高く(図3C)、コラーゲン結合インテグリンがEGFシグナル伝達の阻害に特に関係しているということを示唆している。
【0040】
本発明者らは、ついでEGFRリン酸化のコラーゲン−誘導阻害がTCPTPの活性化を通して媒介されているかどうかの試験を決定した。TCPTPに特異的な低分子干渉RNAs(siRNA)は、HeLa細胞においてTCPTPの内因性発現を効率的に減少させたが、スクランブル対照siRNAは減少させなかった(図3D)。それらはまた、コラーゲン上に置かれた細胞でEGF−誘導EGFRリン酸化を有意に増加させた(図3D)。これらのノックダウン実験は、EGFRシグナル伝達のコラーゲン誘導阻害におけるTCPTPの役割を確認する。
【0041】
HeLa細胞は、α2β1インテグリンと同様にα1β1インテグリンによりコラーゲンに接着できる。インテグリン−TCPTP−EGFRシグナル伝達経路のインテグリン特異性を試験するために、本発明者らは3つのストラテジーを用いた。第1は、α1β1インテグリンを発現しないHT1080線維肉腫細胞において、コラーゲンへの接着はEGFRリン酸化を阻害することはできなかった(図2E)。重要なことは、一時的にα1インテグリンを発現するようにトランスフェクトされたHT1080細胞は、EGFRリン酸化のコラーゲン誘導阻害に感受性となった(図2F)。第2は、接着、血清枯渇HeLa細胞において、モノクローナル抗体とインテグリンα1−サブユニットのクラスタリングは、細胞質PTP活性を1.5倍増加させ、EGFRのEGF−誘導リン酸化を阻止した(図2G)。最終的に、本発明者らは、α1インテグリンヌルマウスおよびその野生型同腹子由来のマウス胚線維芽細胞を用いてその特異性を調べた7。野生型とα1−/−細胞は両方コラーゲンに接着した9(図3E)。しかしながら、α1β1インテグリンの非存在下では、TCPTPは接着部位に見出されず、一方、野生型細胞ではインテグリンα1β1とTCPTPが細胞の周囲の接着部位に同局在化した(図3E)。α1−/−細胞は、コラーゲンまたはフィブロネクチン上に置かれた場合、同等のEGF−誘導EGFRリン酸化を示し、一方、野生型細胞では、コラーゲンへの接着がEGF−誘導シグナル伝達を阻害する結果となった(図3F)。まとめると、これらの実験はα1−インテグリンがTCPTPを特異的に活性化し、この相互作用がEGFRのリン酸化の阻害をもたらすということを確認する。
【0042】
TCPTPの変異体または野生型を用いる過剰発現研究は、細胞増殖の調節におけるその役割を証明した20〜23。さらに、TCPTPは、変異体EGFRを発現するグリア芽腫細胞の増殖および足場非依存性成長を抑制することが示されている23。しかしながら、これまでのところ、TCPTP活性の調節、したがって、その阻害機能のインビボでの実行の機序は知られていない。本発明者らは、インテグリンα1細胞質テイルがインビトロでTCPTPを活性化する能力があることを本明細書に示した。それが細胞においてもホスファターゼ活性を誘導するかどうかを試験するために、本発明者らは、非蛍光ジホスホフルオレセイン(FDP)を、α1またはα2細胞質テイルペプチドと一緒にHeLa細胞中に微量注入した。FDPは脱リン酸化の際に蛍光を発するようになるので、本発明者らは、生HeLa細胞においてホスファターゼ活性を調べた。これらの定量アッセイは、α2またはPBSを同時注入した細胞と比較した場合、インテグリンα1細胞質ペプチドがHeLa細胞においてホスファターゼ活性を誘導したことをはっきりと示した(図4A)。
【0043】
ホスファターゼ活性の誘導は、TCPTPの過剰発現と同様の様式で細胞の腫瘍形成性に影響を与え得る23。最近、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインが、生細胞中にペプチドおよびタンパク質を輸送するために使用されている24、25。したがって、本発明者らは、増殖および腫瘍形成アッセイ用に細胞にα1−cytペプチドを送達するために、TAT−タンパク質に融合した細胞透過性FITC−共役ペプチドを使用した。α1cyt−TATペプチドおよびスクランブル対照ペプチドは両方細胞に効率的に入り(図4B)、蛍光は24時間以上持続し、細胞の剥離は観察されなかった(示さず)。懸濁液中、5%血清で増殖するHeLa細胞では、200nM α1−TATペプチドが血清誘導細胞増殖を阻害した(図4B)。これに対して、対照ペプチドでは増殖は有意に変化しなかった。さらに、α1−TATペプチドはEGF誘導細胞増殖の阻害に有用であるが、懸濁液中の血清不含条件に維持されているHeLa細胞の基礎増殖および生存には効果はなかった(図4D)。腫瘍形成性B104−1−1細胞(neu−腫瘍遺伝子で形質転換された3T3線維芽細胞)の血清誘導増殖もα1−TATペプチドによって阻害された(図4C)。興味深いことに、接着HeLa細胞の増殖は影響されなかったので、α1−TATペプチドは、足場非依存性、EGF−促進(driven)増殖の調節にのみ有効であった(図3G)。さらに、9日間軟寒天で培養されたHeLa細胞による大きな腫瘍コロニーの形成は、α1−TATペプチドによって有意に阻害されたが、対照ペプチドはコロニーサイズを有意に減少させることはできなかった(図4D)。最終的に、インビボでのマウスにおける皮下腫瘍としてのヒト線維肉腫細胞の増殖は、α1−TATペプチドによる処置により阻害された(図5A)。興味深いことに、詳細な免疫組織化学的研究により、α1−TATペプチドで処置した腫瘍は、Scr−TATのPBS注射のいずれかで処置した対照腫瘍よりもより壊死を示すということが明らかとなった(図5B)。これらの実験により、インビボでα1細胞質テイルペプチドによるホスファターゼ活性の誘導が、腫瘍形成性細胞におけるTCPTPの過剰発現により達成される効果と同一の効果をもたらすということが証明される20、23。さらに、ペプチドは生細胞に導入することができ、効率的に悪性細胞の足場非依存性、EGF誘導増殖を阻止し、インビボでの正常細胞増殖の維持におけるα1−送達負シグナルの可能な役割を示唆する。
【0044】
インテグリンのα−細胞質ドメインは、インテグリン媒介生物応答の調節に不可欠であるが26、これまでのところ、ほんの少しの相互作用タンパク質が同定されているのみである。インテグリンは受容体チロシンキナーゼシグナル伝達を正に調節すること、さらにリガンドの非存在下でRTK活性化を誘発することが知られている3、27。また、遍在して発現されるインテグリンα1は、コラーゲンに応答して一次細胞でERK MAP−キナーゼ経路を活性化することが示されている9。他方では、インテグリンα1は、乳癌、卵巣癌および肺腺癌においてダウンレギュレートされると思われる3、27、28。TCPTPのインテグリンα1β1媒介活性化によるEGFRシグナル伝達の負の調節は、細胞応答の接着媒介制御における新規な機序であると思われる。これらの知見は、細胞におけるこの腫瘍抑制タンパク質のホスファターゼ活性の制御に対する分子的説明を提供し、なぜTCPTP(他のPTPsのように)が単離タンパク質としては不規則であるのに、細胞においては高度に特異的であるのかという1つの機序を証明する29。インテグリンα1によるTCPTPの局所調節はそのうえ、タンパク質リン酸化−脱リン酸化平衡の正確な局在化制御が不可欠である他のシグナル伝達経路にとっても重要なようである。たとえば、(インテグリンと結合する27)EGFRと極めて接近した原形質膜でのインテグリンα1細胞質ドメインとの相互作用によるTCPTPの局在的活性化は、厳重に調節された方法で、RTKシグナル伝達を制御するための細胞にとっての強力な方法である。α1細胞質ペプチドの生細胞への送達による、細胞の、接着ではなく足場非依存性増殖の阻害は、形質転換細胞を標的化する新しい手段を提供し得る。結論として、腫瘍抑制タンパク質のインテグリンα1β1媒介活性化によるEGFRのシグナル伝達の負の調節は、細胞応答の接着媒介制御における新規なパラダイムである。
【0045】
本発明の方法は、さまざまな実施態様の形態で組み込まれ得、そのうちのほんのいくつかを本明細書に開示していることは認識されよう。当業者には、他の実施態様が存在し、それは本発明の精神から逸脱しないことが自明である。したがって、記載した実施形態は例示であり、制限的なものと解釈されるべきでない。
【0046】
【図面の簡単な説明】
【0047】
【図1A】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。PC3細胞を、1時間コラーゲン、フィブロネクチンまたはポリ−L−リジンに接着した。インテグリンα1サブユニットおよびTCPTPが二色免疫蛍光染色により検出された。矢印は、膜での同局在化の各領域をそれぞれ指し示す。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して評価した、バー5μm。
【図1B】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞を、1時間コラーゲン、フィブロネクチンまたはポリ−L−リジンに接着した。インテグリンα1サブユニットおよびTCPTPが二色免疫蛍光染色により検出された。矢印は、膜での同局在化の各領域をそれぞれ指し示す。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して評価した、バー5μm。
【図1C】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。血清枯渇HeLa細胞を表面ビオチン化し、プラスチック上に置き、10%FBSで30分間刺激する(FBS)か、コラーゲンI(CI)またはポリ−L−リジン(PL)上に1時間置いた。免疫沈降(ip)されたインテグリン、TCPTPおよび対照(IgG)を免疫ブロッディングで検出した。
【図1D】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞を図1Cと同様に処理し、ライセートを免疫沈降し(ip)、TCPTPに対しブロットするか、または変性させ、抗−α1抗体で再沈降(re−ip)させ、示したように免疫ブロットした。
【図1E】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。HeLa細胞ライセートを固定化グルタチオン−S−トランスフェラーゼ(GST)またはGST−インテグリン細胞質ドメイン融合タンパク質と共にインキュベートした。結合したタンパク質およびライセート試料をTCPTPについてプローブした。ポンソーS染色をローディング対照に使用した。
【図1F】TCPTPがインテグリンα1鎖の細胞質ドメインと結合するということを示す。ポンソーS染色をローディング対照に使用した。組み換えおよび精製TCPTP(1μg)をGSTまたはGST−融合タンパク質と、α1インテグリン細胞質ドメインペプチド(1μg/ml)と共にまたはなしでインキュベートした。結合したタンパク質(およびTCPTPローディング対照、全体)をTCPTPおよびGSTについてプローブした。
【図1G】血清枯渇HeLa細胞を剥離し、コラーゲンまたはポリ−L−リジン上に置き、免疫沈降(ip)にかけ、そしてホスファターゼ活性(平均±SD、n=3)を測定した。試料の半分をTCPTPについてイムノブロットした。
【図1H】血清枯渇HeLa細胞をPBS、対照IgG、または抗−α1 mAbとインキュベートし、抗−マウス二次抗体により30分間クラスタリングを誘導した。ライセート由来の等量のタンパク質のホスファターゼ活性(平均±SD、n=3)を三重で分析した。
【図2A】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。HeLa細胞を、対照(IgG)、抗−TCPTPおよび抗−SHP−2抗体と共に免疫沈降(ip)した。ビヒクル(c)および合成α1細胞質テイルペプチド(α1pep)および合成α2細胞質テイルペプチド(α2pep)での処置後、ホスファターゼ活性(平均±SD、n=3)を、diFMUPを基質として用いて分析した。免疫沈殿物の半分をSDS−PAGE上で分離し、TCPTPまたはSHP−2について標識した。
【図2B】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。組み換え、精製TCPTP(0.15μg/ml)を、異なる濃度のペプチドとインキュベートし、ホスファターゼ活性(平均±SD、n=3)を分析した。
【図2C】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。TCPTP欠失変異体の概略的ダイアグラムを示す。
【図2D】インテグリンα1細胞質テイルがTCPTPを活性化することを示す。競合アッセイでは、全長TCPTPを添加する前に、GSTと融合したTCPTP欠失変異体、またはGSTのみを示したペプチドと共にインキュベートし、ホスファターゼ活性(平均±SD、n=3)を測定した。
【図2E】インテグリンα1β1がEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。FACSによってインテグリンの表面発現をHeLa細胞およびHT1080細胞で分析した。MFI値を示す。血清枯渇HeLa細胞およびHT1080細胞をコラーゲンI(CI)またはフィブロネクチン(FN)上に置き、EGFで処置した。EGFRリン酸化をホスホ特異的抗体を用いて調べた。チューブリンをローディング対照として使用した。
【図2F】インテグリンα1β1はEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。α1−cDNAで一時的にトランスフェクトされたHT1080細胞またはモック−トランスフェクトHT1080細胞から血清を枯渇させ、コラーゲンI(CI)上に置き、EGFで処理した。EGFRリン酸化は図2Eと同様にして分析した。
【図2G】インテグリンα1β1はEGFRシグナル伝達のコラーゲン誘導減衰に必要とされるということを示す。血清枯渇HeLa細胞を図1Hと同様に処理してα1受容体と交差させ、EGFRリン酸化をイムノブロッディングにより調べた。
【図3A】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持された、またはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。チューブリンをローディング対照として使用した。
【図3B】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持されたまたはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。EGFRをローディング対照として使用した。
【図3C】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。プラスチック上に維持されたまたはコラーゲン(CI)もしくはフィブロネクチン上に置かれた血清枯渇HeLa細胞を、EGFで処理し、EGFRリン酸化をホスホ−特異的抗体を用いて調べた。3つ(A、C)の実験のデンシトメトリーによる分析(平均±SD)が示される。チューブリンをローディング対照として使用した。
【図3D】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。TCPTPに対して特異的な2つのsiRNAs(またはスクランブル対照)でトランスフェクトされたHeLa細胞を、コラーゲン上に置き、EGFで処理した。抽出物を示したタンパク質についてイムノブロットした。同様の結果を有する3つの実験の代表を示す。
【図3E】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を減衰させることを示す。α1−/−マウスまたはその野生型同腹子由来の線維芽細胞をコラーゲン上に置き、インテグリンα1およびTCPTPに対して免疫染色した。緑と赤のピクセル強度(平均±SEM、n=12)を、共焦点顕微鏡ソフトウエアを用いて細胞の縁から開始して分析した;バー5μm。
【図3F】インテグリンα1β1のライゲーションが、TCPTPの活性化を通してEGFRリン酸化を軽減することを示す。α1−/−および+/+動物由来の血清枯渇線維芽細胞をコラーゲンまたはフィブロネクチン上に置き、EGFで処理し、イムノブロットした。値はチューブリンに標準化したデンシトメトリーによる評価法である。
【図3G】インテグリンα1細胞質ペプチドがマトリックス接着細胞のEGF非依存性増殖に影響を及ぼさないことを示す。HeLa細胞をコラーゲンまたはフィブロネクチンでコートしたウェル中、5%血清で、200nM TATペプチドおよび50ng/ml EGFの存在下、または非存在下で培養し、種々の時点での生細胞の数(平均±SD、n=3)を分析した。
【図4A】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。HeLa細胞を、脱リン酸化の際に蛍光を発するようになるフルオレセインジホスフェート(FDP)と細胞質インテグリンペプチドと共に微量注入した(アスタリスク)。蛍光強度を30分間モニターし(30分での代表的な画像を示す。)、個々の細胞における平均強度(平均±SEM、n=12)を評価した。
【図4B】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。FITC標識TAT−α1細胞質テイル融合ペプチド(α1−TAT)およびTAT−スクランブル対照融合ペプチド(Scr−TAT)をHeLa細胞に入れたことを示す。5%血清、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での非接着生HeLa細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4C】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。血清不含培地で、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での非接着生HeLa細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4D】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。血清含有培地で、200nM TAT−ペプチドおよび50ng/mlEGFの存在下または非存在下での腫瘍形成性B104−1−1線維芽細胞の数に対するそれらの効果を測定した(平均±SD、n=3)。
【図4E】α1細胞質テイルペプチドがインビボでホスファターゼ活性を誘導し、足場非依存性およびEGF−誘導細胞増殖を阻害するということを示す。HeLa細胞を寒天培地中でTAT−ペプチドと共に、またはなしで9日間増殖させた。代表的な位相差画像を二重盲式で撮影し、コロニーの大きさの分析を示す。
【図5A】α1細胞質テイルペプチドが腫瘍増殖を減少させ、インビボで壊死を誘導することを示す。HT1080ヒト線維肉腫細胞をヌードマウスの側面に注射し、実験項に示したように処理した。皮下腫瘍は4週間後に取り出した。腫瘍の大きさ(領域)を測り、示す(平均±SEM、n=12)。
【図5B】α1細胞質テイルペプチドが腫瘍増殖を減少させ、インビボで壊死を誘導することを示す。HT1080ヒト線維肉腫細胞をヌードマウスの側面に注射し、実験項に示したように処理した。皮下腫瘍は4週間後に取り出した。全腫瘍切断面をマッソン(Masson)トリクロム染色で染色し、全腫瘍領域に対する壊死領域を二重盲式で記録した(平均±SEM、n=12)。壊死領域の分析を示す。
【特許請求の範囲】
【請求項1】
i)アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチド、またはii)哺乳類細胞において該ペプチドを発現可能なベクターのいずれかである、治療における使用のための薬剤。
【請求項2】
前記薬剤がペプチドであって、細胞膜透過融合タンパク質の形態である請求項1記載の薬剤。
【請求項3】
HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項2記載の融合タンパク質。
【請求項4】
個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法での使用のための医薬組成物の製造のための請求項1〜3のいずれか1項に記載の薬剤の使用。
【請求項5】
チロシンキナーゼシグナル伝達の阻害方法、または個体においてチロシンキナーゼシグナル伝達を阻害することによって治癒可能な疾患または障害の予防または治療方法での使用のための医薬組成物の製造のための、該個体においてT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化できる薬剤の使用。
【請求項6】
前記薬剤が請求項1〜3に記載された薬剤のいずれか1つである請求項5記載の使用。
【請求項7】
前記チロシンキナーゼが上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項5または6記載の使用。
【請求項8】
前記疾患が癌である請求項5〜7のいずれか1項に記載の使用。
【請求項9】
癌の予防方法、または個体において癌の成長、浸潤または転移の予防または阻害方法での使用のための医薬組成物の製造のための、該個体においてT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化できる薬剤の使用。
【請求項10】
薬剤が請求項1〜3に記載された薬剤のいずれか1つである請求項9記載の使用。
【請求項11】
治療的有効量のTCPTPを活性化できる低分子またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれか、および薬学的に許容され得る担体を含む医薬組成物。
【請求項12】
前記アミノ酸配列RPLKKKMEK(配列番号1)が、細胞膜透過性である融合タンパク質に包含される請求項11記載の組成物。
【請求項13】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインとアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項12記載の組成物。
【請求項14】
前記担体が組成物の非経口投与に適した請求項11〜13のいずれか1項に記載の組成物。
【請求項15】
アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドをコードする核酸を包含する発現ベクターであり、哺乳動物細胞において該ペプチドを発現することができるベクター、および薬学的に許容され得る担体を含む医薬組成物。
【請求項16】
核酸がDNA配列、RNA配列またはウイルスベクター中に挿入されている請求項15記載の組成物。
【請求項17】
ウイルスベクターがアデノウイルス、アルファウイルス、アデノ随伴ウイルス、レトロウイルスまたはヘルペスウイルスに基づくものである請求項16記載の組成物。
【請求項18】
前記発現ベクターが、脂質と複合化されるか、リポソームに詰められているか、シクロデキストリンもしくは他の複合化剤、生吸収性ポリマーまたは制御された放出投与のための他の好適な担体に組み込まれているか、ナノ粒子またはヒドロゲルに包含されている請求項15〜17のいずれか1項に記載の組成物。
【請求項19】
個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法であって、該個体に、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである有効量の薬剤を投与することによる方法。
【請求項20】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項19記載の方法。
【請求項21】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項20記載の方法。
【請求項22】
個体におけるチロシンキナーゼシグナル伝達を阻害する方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項23】
前記チロシンキナーゼが、上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項22記載の方法。
【請求項24】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項22または23記載の方法。
【請求項25】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項24記載の方法。
【請求項26】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項25記載の方法。
【請求項27】
個体におけるチロシンキナーゼシグナル伝達を阻害することにより治癒可能な疾患または障害の予防または治療方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項28】
前記チロシンキナーゼが、上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項27記載の方法。
【請求項29】
前記疾患が癌である請求項27または28記載の方法。
【請求項30】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項27〜29のいずれか1項に記載の方法。
【請求項31】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項30記載の方法。
【請求項32】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項31記載の方法。
【請求項33】
個体における、癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項34】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項33項記載の方法。
【請求項35】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項34記載の方法。
【請求項36】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項35記載の方法。
【請求項1】
i)アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチド、またはii)哺乳類細胞において該ペプチドを発現可能なベクターのいずれかである、治療における使用のための薬剤。
【請求項2】
前記薬剤がペプチドであって、細胞膜透過融合タンパク質の形態である請求項1記載の薬剤。
【請求項3】
HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項2記載の融合タンパク質。
【請求項4】
個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法での使用のための医薬組成物の製造のための請求項1〜3のいずれか1項に記載の薬剤の使用。
【請求項5】
チロシンキナーゼシグナル伝達の阻害方法、または個体においてチロシンキナーゼシグナル伝達を阻害することによって治癒可能な疾患または障害の予防または治療方法での使用のための医薬組成物の製造のための、該個体においてT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化できる薬剤の使用。
【請求項6】
前記薬剤が請求項1〜3に記載された薬剤のいずれか1つである請求項5記載の使用。
【請求項7】
前記チロシンキナーゼが上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項5または6記載の使用。
【請求項8】
前記疾患が癌である請求項5〜7のいずれか1項に記載の使用。
【請求項9】
癌の予防方法、または個体において癌の成長、浸潤または転移の予防または阻害方法での使用のための医薬組成物の製造のための、該個体においてT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化できる薬剤の使用。
【請求項10】
薬剤が請求項1〜3に記載された薬剤のいずれか1つである請求項9記載の使用。
【請求項11】
治療的有効量のTCPTPを活性化できる低分子またはアミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドのいずれか、および薬学的に許容され得る担体を含む医薬組成物。
【請求項12】
前記アミノ酸配列RPLKKKMEK(配列番号1)が、細胞膜透過性である融合タンパク質に包含される請求項11記載の組成物。
【請求項13】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインとアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項12記載の組成物。
【請求項14】
前記担体が組成物の非経口投与に適した請求項11〜13のいずれか1項に記載の組成物。
【請求項15】
アミノ酸配列RPLKKKMEK(配列番号1)を含むペプチドをコードする核酸を包含する発現ベクターであり、哺乳動物細胞において該ペプチドを発現することができるベクター、および薬学的に許容され得る担体を含む医薬組成物。
【請求項16】
核酸がDNA配列、RNA配列またはウイルスベクター中に挿入されている請求項15記載の組成物。
【請求項17】
ウイルスベクターがアデノウイルス、アルファウイルス、アデノ随伴ウイルス、レトロウイルスまたはヘルペスウイルスに基づくものである請求項16記載の組成物。
【請求項18】
前記発現ベクターが、脂質と複合化されるか、リポソームに詰められているか、シクロデキストリンもしくは他の複合化剤、生吸収性ポリマーまたは制御された放出投与のための他の好適な担体に組み込まれているか、ナノ粒子またはヒドロゲルに包含されている請求項15〜17のいずれか1項に記載の組成物。
【請求項19】
個体におけるT細胞タンパク質チロシンホスファターゼ(TCPTP)の活性化方法であって、該個体に、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである有効量の薬剤を投与することによる方法。
【請求項20】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項19記載の方法。
【請求項21】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項20記載の方法。
【請求項22】
個体におけるチロシンキナーゼシグナル伝達を阻害する方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項23】
前記チロシンキナーゼが、上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項22記載の方法。
【請求項24】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項22または23記載の方法。
【請求項25】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項24記載の方法。
【請求項26】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項25記載の方法。
【請求項27】
個体におけるチロシンキナーゼシグナル伝達を阻害することにより治癒可能な疾患または障害の予防または治療方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項28】
前記チロシンキナーゼが、上皮成長因子受容体(EGFR)、インスリン受容体およびヤーヌスキナーゼからなる群より選択される請求項27記載の方法。
【請求項29】
前記疾患が癌である請求項27または28記載の方法。
【請求項30】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項27〜29のいずれか1項に記載の方法。
【請求項31】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項30記載の方法。
【請求項32】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項31記載の方法。
【請求項33】
個体における、癌の予防、または癌の成長、浸潤もしくは転移の予防または阻害方法であって、該個体にT細胞タンパク質チロシンホスファターゼ(TCPTP)を活性化する能力を有する薬剤を有効量投与することによる方法。
【請求項34】
前記薬剤が、i)アミノ酸配列RPLKKKMEK(配列番号1)またはii)哺乳動物細胞において該ペプチドを発現することができるベクターのいずれかである請求項33項記載の方法。
【請求項35】
前記薬剤がペプチドであり、それ自体として投与されるかまたは細胞膜透過融合タンパク質として投与される請求項34記載の方法。
【請求項36】
前記融合タンパク質が、HIV転写トランスアクチベーター(TAT)タンパク質のタンパク質形質導入ドメインおよびアミノ酸配列RPLKKKMEK(配列番号1)を含む請求項35記載の方法。
【図1A】

【図1B】
【図1C】
【図1D】
【図1E】
【図1F】

【図1G】
【図1H】
【図2A】

【図2B】
【図2C】
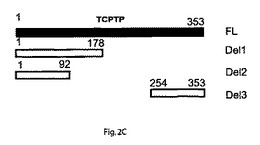
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】

【図3F】
【図3G】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】

【図5A】
【図5B】

【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図1G】
【図1H】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図3G】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図5A】
【図5B】
【公表番号】特表2008−509085(P2008−509085A)
【公表日】平成20年3月27日(2008.3.27)
【国際特許分類】
【出願番号】特願2007−513979(P2007−513979)
【出願日】平成17年5月2日(2005.5.2)
【国際出願番号】PCT/FI2005/000203
【国際公開番号】WO2005/118639
【国際公開日】平成17年12月15日(2005.12.15)
【出願人】(501374390)バルティオン テクニリーネン トゥトキムスケスクス (16)
【Fターム(参考)】
【公表日】平成20年3月27日(2008.3.27)
【国際特許分類】
【出願日】平成17年5月2日(2005.5.2)
【国際出願番号】PCT/FI2005/000203
【国際公開番号】WO2005/118639
【国際公開日】平成17年12月15日(2005.12.15)
【出願人】(501374390)バルティオン テクニリーネン トゥトキムスケスクス (16)
【Fターム(参考)】
[ Back to top ]