
Fターム[2B030CB02]の内容
植物の育種及び培養による繁殖 (7,935) | 実施例、実験例の開示(分化の確認等) (1,021) | 分化(胚、不定芽等)の確認ができるもの (896) | 植物体再生の確認ができるもの (796)
Fターム[2B030CB02]に分類される特許
161 - 180 / 796
遺伝的に改変された細胞を製造する方法
【課題】二重鎖切断誘導による相同組み換えを利用して遺伝子的に改変された細胞を製造する手法において、二重鎖切断部位における相同組み換えの頻度を高めること。
【解決手段】ブルームDNAヘリカーゼおよび/またはエキソヌクレアーゼExo1の利用により、二重鎖切断誘導による相同組み換えの頻度を飛躍的に高めることができることを見出した。
(もっと読む)
倍加半数体植物を作出する方法
本発明は、倍加半数体植物を作出する方法であって、機能的精細胞を一つ有する花粉を中央細胞ではない胚嚢細胞と受精させる工程; 中央細胞を増殖させて胚乳とする工程; および該胚乳から倍加半数体植物を再生する工程を含む方法に関する。機能的精細胞を一つ有する花粉は、例えば、化学的突然変異、核酸を用いる形質転換または照射によって得ることができる突然変異体花粉である。 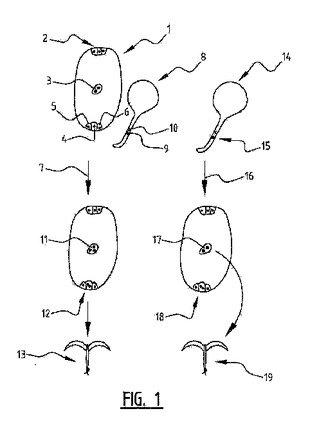 (もっと読む)
(もっと読む)
ポリ不飽和脂肪酸に富む油を産生するための新規Δ9−エロンガーゼ
本発明は、Δ9−エロンガーゼをコードする単離ポリヌクレオチド、前記単離ポリヌクレオチドによりコードされるΔ9−エロンガーゼ、前記単離ポリヌクレオチドを含む発現ベクター、前記発現ベクターを含む宿主細胞、並びにΔ9−エロンガーゼ及びポリ不飽和脂肪酸の産生方法に関する。  (もっと読む)
(もっと読む)
病原体抵抗性の植物
本発明は、ブレミア抵抗性の新規ラクツカ・サティバ(Lactuca sativa)植物、および前記植物の種子に関する本発明はまた、そのような植物およびその種子を作製する方法に関する。本発明はさらに、マーカー利用による育種における、ならびにブレミア抵抗性形質を同定するための、マーカーおよびその方法に関する。 (もっと読む)
植物および真菌において2−アセチル−1−ピロリン合成を増強する核酸
【課題】芳香族化合物2−アセチル−1−ピロリン(2AP)は、あらゆる香米の主要な強い香気成分であり、正確な生合成経路はまだ解明されていない2−アセチル−1−ピロリン合成に関与する遺伝子の同定、さらに植物および真菌において2−アセチル−1−ピロリンレベルを高めて芳香を増強するための方法を提供する。
【解決手段】2−アセチル−1−ピロリンが、天然に存在する非芳香性種で合成されるレベルよりも高いレベルで合成される非天然植物。さらに、より高いレベルの2−アセチル−1−ピロリンを合成する植物および真菌を選択するための分子マーカーとして使用することができる核酸。
(もっと読む)
植物細胞へのDNAの導入

本発明は、外来性遺伝物質を植物細胞に簡便かつ効率的に導入するための手段および方法を提供する。特に、本発明は、植物中への異種DNAの効率的な導入のために、種子プライミングとウイルスに基づくDNA構築物を組み合わせる。 (もっと読む)
グリホサート耐性が増強された植物を得るための方法および手段
本発明は、植物の構成的プロモーターの制御下のグリホサート耐性EPSPS酵素と、置換ヒストンイントロン1とをコードするキメラDNA分子を有し、それによって、増強されたグリホサート耐性を前記植物に付与する、植物に関する。  (もっと読む)
(もっと読む)
トマトにおける受精非依存果実形成
本発明は、受精非依存果実形成の形質を含むトマト植物に関し、該形質は、その代表種子がNCIMBに受託番号 NCIMB 41626、NCIMB 41627、NCIMB 41628、NCIMB 41629、NCIMB 41630またはNCIMB 41631にて寄託されている植物からの遺伝子移入によって得られうるものである。かかるトマト植物は、その代表種子がNCIMB に受託番号 NCIMB 41626、NCIMB 41627、NCIMB 41628、NCIMB 41629、NCIMB 41630またはNCIMB 41631にて寄託されている植物と、該形質を示さない植物とを交配してF1 集団を得る工程; F1 集団からの植物を自家受粉させて F2 集団を得る工程; F2 植物の授粉を阻止して果実形成が起こることを可能にする工程;および、果実を生産する植物を受精非依存果実形成を示す植物として選抜する工程によって得られうる。本発明はさらに、単為結実果実、該植物の種子および該植物の繁殖材料にも関する。 (もっと読む)
種子生産性が安定化された高妊性ラパノブラシカ新品種植物

本発明は、種子生産性が安定化された高妊性ラパノブラシカ新品種植物の育種方法に係り、突然変異誘発物質NMUを0.01μg/L処理してラパノブラシカ種子を突然変異処理して種まきして採種してまた種まき後室内で人工交配して種子生産性を確認して続いてAFLPプライマー(primer)を利用して品種の均一性を確認することを特徴とするラパノブラシカ新品種植物の育種方法を開示する。 (もっと読む)
ヒドロラーゼ、それをコードする核酸並びにその製造および使用方法
【課題】本発明は、ヒドロラーゼ、それをコードするポリヌクレオチド、並びにこれらのポリヌクレオチドおよびポリペプチドを製造および使用する方法を提供する。
【解決手段】ある特徴では、本発明は、ポリペプチド(例えばヒドロラーゼ活性(例えばエステラーゼ、アシラーゼ、リパーゼ、ホスホリパーゼ(例えばホスホリパーゼA、B、CおよびD活性、パタチン活性、脂質アシルヒドロラーゼ(LAH)活性)、またはプロテアーゼ活性であり、熱安定性および耐熱性のヒドロラーゼ活性を含む)を有する酵素)、およびこれらの酵素をコードするポリヌクレオチド、並びに、これらのポリヌクレオチドおよびポリペプチドの製造および使用を対象とする。
(もっと読む)
DIG−10殺虫性CRY毒素
DIG−10Cry毒素、かかる毒素をコードするポリヌクレオチド、有害生物を防除するためのかかる毒素の使用、及びかかる毒素を産生する遺伝子組換え植物が開示される。 (もっと読む)
キシロシルトランスフェラーゼ活性の欠損したベンサミアナタバコ(Nicotianabenthamiana)植物
本発明は、糖タンパク質上でキシロシル構造を形成できないベンサミアナタバコ(Nicotiana benthamiana)の突然変異植物を提供する。加えて本発明は、前記突然変異植物における異種糖タンパク質の産生のための方法を提供する。  (もっと読む)
(もっと読む)
DIG−5殺虫性CRY毒素
DIG−5Cry毒素、かかる毒素をコードするポリヌクレオチド、有害生物を防除するためのかかる毒素の使用、及びかかる毒素を産生する遺伝子組換え植物が開示される。 (もっと読む)
種子における物質生産性を向上させる遺伝子及びその利用方法
【課題】物質生産性、特に種子における油脂含量を増加或いは減少させることができる新規な機能を有する遺伝子の提供。
【解決手段】特定のアミノ酸配列を含むタンパク質からなる転写因子と、任意の転写因子を転写抑制因子に転換する特定のアミノ酸配列を含む機能性ペプチドとを融合させたキメラタンパク質。該キメラタンパク質を発現させた植物体。該キメラタンパク質をコードする遺伝子。該遺伝子を含む発現ベクター。該遺伝子を含む形質転換体。
(もっと読む)
種子のタンパク質含量を増産させる遺伝子及びその利用方法
【課題】種子におけるタンパク質含量を増加或いは減少させることができる新規な機能を有する遺伝子を提供する。
【解決手段】特定の配列のうちいずれか1つのアミノ酸配列を含むタンパク質からなる転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を植物体において発現させる。若しくは特定の配列のうちいずれか1つのアミノ酸配列を含むタンパク質からなる転写因子を植物体において過剰発現させる。
(もっと読む)
植物の植物重量を増産させる遺伝子及びその利用方法
【課題】植物重量すなわちバイオマス量を増産させるとともに物質生産性を増加或いは減少させることができる新規な機能を有する遺伝子を探索する。
【解決手段】特定のアミノ酸配列からなる転写因子と、任意の転写因子を転写抑制因子に転換する機能性ペプチドとを融合させたキメラタンパク質を発現させた植物体。植物体がアブラナ科植物のシロイヌナズナであり、該植物の重量、種子に含まれる油脂の生産性が有意に向上又は減少した形質転換体。
(もっと読む)
植物に除草剤抵抗性を付与するための遺伝子およびベクター
【課題】イミダゾリノンなどの除草剤に対する耐性または抵抗性レベルの上昇したトランスジェニック植物を提供する。
【解決手段】真核生物のAHAS小サブユニットタンパク質をコードする、ゲノムおよびcDNA配列並びに植物発現ベクターを使用して、植物を形質転換する。
(もっと読む)
非結球性メキャベツ類及びその作成方法
【構成】 メキャベツの純系を種子親、アセファラGP1の純系を花粉親として交配し、カルシム含有量が2,880 mg/100g以上とした非結球性メキャベツ類(請求項1)である。
アセファラGP1の純系を種子親、アセファラGP2の純系を花粉親として交配し、βカロチン含有量が60mg/100g以上であるようにした非結球性メキャベツ類(請求項3)である。
【効果】 表2にに示すような栄養価の高い非結球性メキャベツ類を提供することができる。生鮮野菜として、また、健康補助食品(青汁等)の原料として、栄養価の高い非結球性メキャベツ類を利用することができる。
(もっと読む)
新規糖トランスポーター
SWEET、GLUEまたはGlueと称するトランスポータータンパク質の新規クラスを開示する。これらのトランスポーターは、細胞内および細胞の内と外の間で膜を通過する糖輸送の新規システムを提供する。かかるトランスポーターは、生物の特定の臓器および細胞の特定の細胞小器官内の糖濃度を理解し、変化するために有用である。これらのトランスポーターはまた、病原体侵襲からの植物の保護にも有用である。 (もっと読む)
植物の生長を制御する遺伝子Hd16およびその利用
【課題】本発明は、植物の生長を制御する新規遺伝子の提供、該遺伝子を利用した植物の生長の制御(開花期又は出穂期の改変)方法、及び該遺伝子を標的とした植物の感光性の強度の判定方法を提供することを課題とする。
【解決手段】本発明者らは、上記の課題を解決するために、イネ品種日本晴とコシヒカリの間で検出された植物の生長を制御する遺伝子座(Hd16遺伝子座)の高精度連鎖解析を行い、Hd16の候補遺伝子を選定した。さらに、上記のようにして絞り込んだ候補遺伝子が、実際に出穂に関して機能していることを確認するために相補性検定を行った。その結果、Hd16候補遺伝子として単離した遺伝子には到穂日数を調節する機能を有することが明らかとなり、さらに該当遺伝子を使用して出穂期および植物の生長を制御することが可能であることを見出した。
(もっと読む)
161 - 180 / 796
[ Back to top ]