
独立行政法人農業生物資源研究所により出願された特許
11 - 20 / 200
病害ストレス及び病害抵抗性誘導剤に対する応答性を示す核酸構築物
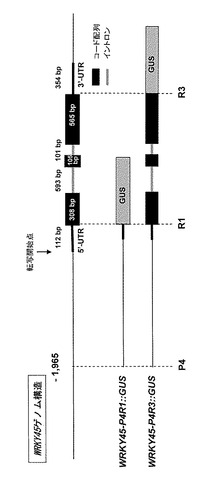
【課題】イネにおけるWRKY45遺伝子の応答性を明らかにし、病害ストレス及び病害抵抗性誘導剤に対する応答性を示す新規な核酸構築物を提供する。
【解決手段】本発明に係る核酸構築物は、配列番号1の塩基配列からなる、イネにおけるWRKY45遺伝子の転写開始点上流1965から3'-UTRの直前までの領域からなるポリヌクレオチドと、目的タンパク質をコードするポリヌクレオチドとを連結してなる。
(もっと読む)
植物のtan遺伝子およびその用途
【課題】 植物の病害虫等による外的障害に対する色素反応に関わり、病原性糸状菌に対する感受性および抵抗性を支配する遺伝子を同定すること。
【解決手段】 病原性糸状菌に対する抵抗性品種と感受性品種とを交配させて得られたF3からF5の集団を対象として、3-デオキシアントシアニジンの合成を担い、ソルガムに当該感受性をもたらすtan遺伝子のマッピングを行った結果、当該遺伝子の同定に成功した。さらに、tan遺伝子がコードする蛋白質の機能喪失により、ソルガムに当該抵抗性が付与されることを見出した。
(もっと読む)
青枯病抵抗性誘導剤及び青枯病防除方法

【課題】青枯病に対する抵抗性誘導活性を示す化合物を有効成分として含有してなる青枯病抵抗性誘導剤及び該化合物を利用した青枯病防除方法を提供する。
【解決手段】L体のアミノ酸を有効成分として含有する青枯病抵抗性誘導剤。L体のアミノ酸を、トマトなどの対象植物に吸収させることによって、青枯病を防除することができる。特にL体のアミノ酸が、L−ヒスチジン、L−アルギニン、L−リシン、L−アスパラギン酸、L−グリシン、L−システイン、L−フェニルアラニン、L−プロリン、L−アラニン、L−グルタミン及びL−メチオニンからなる群から選ばれる少なくとも1種であることが好ましい。
(もっと読む)
カイコの卵および眼の着色に関与する遺伝子およびその利用
【課題】 カイコのre遺伝子を同定すること、同定したre遺伝子を利用して、カイコの卵および眼の着色を改変すること、および、同定したre遺伝子を遺伝子マーカーとして利用して、遺伝子組換えカイコを選抜すること
【解決手段】 ポジショナルクローニングの手法などを利用して、re遺伝子を同定し、カイコの卵および眼における赤色の着色が、この遺伝子の機能が喪失によって標準型の色素が合成されないことにより生じていることを見出した。当該遺伝子を利用することにより、カイコの卵および眼の色の改変を行うことが可能であり、さらには、このような形質の変化を指標として、効率的に遺伝子組換えカイコを選抜することが可能であることを見出した。
(もっと読む)
植物変異体、植物変異体の製造方法、及び可溶糖の蓄積方法
【課題】植物の種子以外の組織において、可溶糖を高蓄積する。
【解決手段】植物におけるADP-グルコースピロホスホリラーゼを構成するサブユニットのうち、イネにおけるAGPL1遺伝子がコードするサブユニットと最も配列類似性の高いサブユニットをコードする遺伝子を抑制するように改変した。
(もっと読む)
センチュウ抵抗性誘導剤及びセンチュウ防除方法
【課題】植物抵抗性誘導活性を示す化合物を有効成分として含有してなるセンチュウ抵抗性誘導剤および該化合物を利用したセンチュウ防除方法を提供する。
【解決手段】スクラレオールを有効成分として含有するセンチュウ抵抗性誘導剤。スクラレオール自体は、抗センチュウ活性を有さないが、トマトなどの対象植物にスクラレオールを吸収させることによって、センチュウを防除することができる。
(もっと読む)
カイコ中部絹糸腺特異的遺伝子発現系を利用したタンパク質の製造方法
【課題】カイコの中部絹糸線における組換えタンパク質の生産方法を提供することを課題とする。
【解決手段】セリシン遺伝子のプロモーターによって発現が制御されるGFPを有するトランスジェニックカイコを作出した。該カイコの最終齢の幼虫の絹糸腺を観察した結果、中部絹糸腺でのみ蛍光が観察された。また、吐糸期ころからGFPは中部絹糸腺の細胞から分泌され、GFPが腺腔内に移動していることがわかった。最終的にはGFPは繭糸として吐糸され、GFPを大量に含む繭が作られた。このことから、セリシン遺伝子のプロモーター領域を利用することにより、中部絹糸腺において組換えタンパク質を生産することが可能であることが分かった。また、中部絹糸腺で生産された組換えタンパク質は容易に中部絹糸腺の内腔に分泌されることが分かった。
(もっと読む)
植物の種子休眠を制御するSdr4遺伝子およびその利用
【課題】本発明は、植物の種子休眠を制御する新規な遺伝子を提供することを課題とする。また、該遺伝子を利用して植物の種子休眠を制御する方法を提供することを課題とする。さらに、被検植物の穂発芽耐性を検出する方法を提供することを課題とする。
【解決手段】本発明者らは、上記の課題を解決するために、まず、マップベースクローニング法を利用して、穂発芽耐性遺伝子であるSdr4の塩基配列を単離・同定することに成功した。さらに、相補試験、ノックダウン系統による機能解析、ミュータントの単離により日本晴のSdr4にも発芽抑制能があることを確認した。また、イネSdr4遺伝子と相同性の高いシロイヌナズナの遺伝子を検索した結果、最も相同性の高いAtSdr4L1は、発芽を制御する機能を有することを見出した。
(もっと読む)
種子特異的プロモーターおよびその利用
【課題】種子に特異的活性を有するプロモーターおよび種子に外来タンパク質を発現させる方法を提供することを課題とする。
【解決手段】複数のイネ種子発現遺伝子のプロモーターを単離し、GUSレポーター遺伝子の上流に各プロモーターを挿入したバイナリーベクターをそれぞれ作製し、アグロバクテリウム法によりイネを形質転換した。GUS発現量を指標として、各プロモーターによる発現部位、種子成熟過程での発現、さらに種子での発現強度を検討したところ、種子の特定部位に特異的発現活性を有し、恒常的プロモーターや既知の種子特異的プロモーターと比較して高い活性をもつプロモーターを見出した。
(もっと読む)
植物の種子休眠性を支配する遺伝子およびその利用
【課題】 植物における種子休眠性の程度を支配する新規な遺伝子を同定すること、並びに、同定された遺伝子を利用して、植物の種子休眠性の程度を効率的に判定する方法および種子休眠性が改変された植物を効率的に作出する方法を提供すること
【解決手段】 ポジショナルクローニングの手法により、オオムギの種子休眠性を支配するSD2遺伝子、およびこれに対応するコムギ遺伝子であるPhs1遺伝子を同定することに成功した。さらに、これら遺伝子において、その種子休眠性と相関する一塩基多型(SNP)を特定するとともに、弱種子休眠性型の遺伝子が優性であることを解明することにも成功した。同定した遺伝子を利用して、効率的に、植物の種子休眠性の程度を判定することや種子休眠性が改変された植物を作出することが可能であることを見出した。
(もっと読む)
11 - 20 / 200
[ Back to top ]